
Euoplocephalus
| Euoplocephalus Status: Uitgestorven, als fossiel bekend | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||||
| ROM 1930 | |||||||||||||||||||
| Taxonomische indeling | |||||||||||||||||||
| |||||||||||||||||||
| Geslacht | |||||||||||||||||||
| Euoplocephalus Lambe, 1910 | |||||||||||||||||||
| Typesoort | |||||||||||||||||||
| Stereocephalus tutus Lambe 1902 | |||||||||||||||||||
| Soorten | |||||||||||||||||||
| |||||||||||||||||||
| Afbeeldingen op | |||||||||||||||||||
| Euoplocephalus op | |||||||||||||||||||
| |||||||||||||||||||
Euoplocephalus[1][2] is een geslacht van uitgestorven plantenetende en zwaar bepantserde ornithischische dinosauriërs, behorende tot de Ankylosauria, dat tijdens het Laat-Krijt leefde in het gebied van het huidige Noord-Amerika.
De eerste fossielen van Euoplocephalus zijn in 1897 gevonden in Canada. In 1902 werden deze benoemd als de typesoort Stereocephalus tutus, waarvan de geslachtsnaam echter al bezet bleek door een kever. Daarom is het geslacht in 1910 hernoemd tot Euoplocephalus, wat zoiets als 'welgewapend hoofd' betekent. In het begin van de twintigste eeuw zijn nog veel meer van dit soort fossielen in Noord-Amerika gevonden en vaak benoemd als aparte geslachten. Op het eind van de eeuw wees men die vondsten allemaal aan Euoplocephalus toe, die zo als een van de best gekende dinosauriërs beschouwd werd. Tegenwoordig denken veel wetenschappers echter weer dat het toch om verschillende geslachten ging. Een half dozijn tamelijk volledige skeletten bleef van de echte Euoplocephalus over, zodat de geleerden desalniettemin een goed beeld hebben van de bouw van het dier.
Euoplocephalus werd vijf tot zes meter lang en ruim twee ton zwaar. Hij had een zeer platte schijfvormige romp die rustte op vier korte stevige poten aan de uiterste voorkant en achterkant. Een dikke lange nek droeg een stompe naar beneden gekromde kop. Met een brede hoornsnavel werden allerlei planten afgebeten om in de enorme buikholte verteerd te worden. Net als andere leden van de Ankylosauridae verdedigde Euoplocephalus zich met een benige knots op het einde van de staart tegen roofsauriërs als Gorgosaurus. Zijn lichaam en vooral de kop werd verder beschermd door een groot aantal kleine beenschubben. Rijen grote ovale beenplaten met bovenop een scherpe snijrand liepen over nek, romp en staartbasis naar achteren. De nek was ook omgeven door twee beenringen.
Vondst en naamgeving
[bewerken | brontekst bewerken]Op 18 augustus 1897 vond paleontoloog Lawrence Morris Lambe het eerste fossiel van Euoplocephalus in het gebied van het huidige Dinosaur Provincial Park in Alberta. De precieze locatie is tegenwoordig niet meer bekend, maar werd indertijd aangegeven als de oostelijke oever van de Red Deer River waar de Berry Creek erin uitmondt. In 1902 benoemde Lambe de vondst als Stereocephalus tutus. De geslachtsnaam was afgeleid van het Oudgriekse στερεός, stereos, 'solide', en κεφαλή, kephalè, 'hoofd', een verwijzing naar de bepantsering op de kop. De soortaanduiding betekent 'veilig beschermd' in het Latijn en verwijst opnieuw naar het pantser. De geslachtsnaam was echter al bezet door de kever Stereocephalus Lynch 1884 en moest dus vervangen worden om een geldige soortnaam op te leveren. In 1910 koos Lambe hiervoor de naam Euoplocephalus die is afgeleid van het Griekse eu, 'goed', en hoplo~, een woordelement dat in combinatie gebruikt wordt om een bewapening aan te geven. De naam betekent dus min of meer 'welgewapend hoofd'. Euoplocephalus tutus is daarmee de combinatio nova, de nieuwe combinatienaam, voor de typesoort van het geslacht, die Stereocephalus tutus blijft.

Het holotype CMN 210, eerder NMC 0210, is gevonden in een laag van de onderste Dinosaur Park Formation, die dateert uit het Campanien en ongeveer 76 miljoen jaar oud is. Het bestaat uit een schedel, een stuk van de voorste halsberg, zijnde een van de pantserringen om de nek, en een, nooit geprepareerde, rechteronderkaak. Een rib die vermoedelijk van hetzelfde individu afkomstig is, heeft het aparte inventarisnummer CMN 1463.

Uit het Campanien van Canada zouden daarna nog veel meer ankylosauride vondsten gedaan worden. Vaak werden die benoemd als aparte soorten. Lambe zelf bijvoorbeeld had in dezelfde publicatie waarin hij Stereocephalus beschreef een tand, specimen NMC 1349, benoemd als Palaeoscincus asper. In 1924 benoemde William Arthur Parks een Dyoplosaurus acutosquameus. In 1928 benoemde Franz Nopcsa een Scolosaurus cutleri. In 1929 benoemde Charles Mortram Sternberg een Anodontosaurus lambei.
Sommige wetenschappers meenden dat al die variatie maar schijn was en dat zulke soorten samengevoegd konden worden. In 1915 vond Edwin Hennig dat het geslacht Palaeoscincus identiek was aan Euoplocephalus en Euoplocephalus tutus dus een Palaeoscincus tutus moest zijn. In 1964 bracht Oskar Kuhn Euoplocephalus bij Ankylosaurus onder als een Ankylosaurus tutus. Deze twee namen bleven slechts curiosa waarvan het bestaan nauwelijks doordrong. In 1971 echter promoveerde Walter Preston Coombs op een zeer invloedrijke dissertatie waarin hij onder meer stelde dat al deze vormen identiek waren aan, en dus jongere synoniemen van, Euoplocephalus tutus. Zelfs de Aziatische Tarchia gigantea Maleev 1956 zou hij in 1978 hernoemen tot een tweede soort van Euoplocephalus: Euoplocephalus giganteus. In 2003 werd Coombs standpunt dat al het Noord-Amerikaanse ankylosauride materiaal slechts één soort vertegenwoordigde, Euoplocephalus tutus, nog verdedigd door Matthew Vickaryous.
In het begin van de eenentwintigste eeuw begon echter grote twijfel te ontstaan over de mogelijkheid dat één enkele soort zo'n groot bereik in ruimte en tijd zou hebben kunnen gehad. Door nieuwe vondsten en verbeterde inzichten en technieken werd het ook eenvoudiger het omvangrijke fossiele materiaal te dateren en te groeperen. Dat leidde ertoe dat Anodontosaurus en Scolosaurus weer als geldige taxa werden gezien. Daarbij werd er nog een geslacht benoemd: Oohkotokia. Tarchia was men eigenlijk altijd al als een apart geslacht blijven beschouwen.
Binnen de opvatting van Coombs was Euoplocephalus tutus verreweg de best bekende ankylosauride, waarvan het materiaal bestond uit ruim veertig specimina uit Alberta, Canada en Montana, Verenigde Staten van Amerika. De nieuwe interpretaties beperken de met zekerheid tot de soort behorende fossielen aanzienlijk, hoewel ze nog steeds de meerderheid van de Canadese vondsten vertegenwoordigt. Victoria Arbour stelde in 2013 bij een herziening van het geslacht een lijst op met toe te wijzen specimina. Daarbij verwijderde ze al het materiaal uit de V.S. en beperkte het voorkomen tot de onderste dertig meter van de afzettingen van de Dinosaur Park Formation. Over bleven toen: de specimina AMNH 5337, AMNH 5403, AMNH 5404, AMNH 5405, ROM 1930 en UALVP 31, alle gedeeltelijke skeletten met schedel; AMNH 5406, een gedeeltelijk skelet zonder schedel; CMN 842, een halsberg; CMN 8876, een schedel, TMP 1979.14.74, een fragmentarische schedel en UALVP 47977, een stuk schedeldak. Eerder is vaak gesteld dat van Euoplocephalus alle skeletelementen ontdekt waren, maar de beperking van het materiaal leidt tot hiaten in onze kennis: zelfs bij elkaar gevoegd levert het nu geen complete nek, rug of staart op en van de voet zijn alleen wat losse botten bekend. ROM 1930, in 1914 gevonden door George Fryer Sternberg, is de beste informatiebron aangaande het lichaamspantser met osteodermen in de oorspronkelijke positie en zelfs huidafdrukken. Ook dit specimen is op dit punt echter niet volledig.
In 2018 benoemde Paul Penkalski AMNH 5337 als een apart geslacht Platypelta waaraan hij ook AMNH 5403 toewees. AMNH 5404 wees hij toe aan Scolosaurus cutleri en ROM 1930 maakte hij een Scolosaurus thronus. AMNH 5405 was volgens hem een Scolosaurus sp. Zo bleven AMNH 5406, NHMUK R4947 en UA 31 over waaraan hij schedel TMP 1991.127.1 toevoegde.
Beschrijving
[bewerken | brontekst bewerken]Grootte, algemene bouw en onderscheidende kenmerken
[bewerken | brontekst bewerken]
Euoplocephalus is een middelgrote ankylosauride met een lengte van vijf tot zes meter en een gewicht van ruim twee ton. Enkel de Noord-Amerikaanse Ankylosaurus en Cedarpelta zijn overigens bewijsbaar groter.
Euoplocephalus liep op vier poten. Zijn lichaamsbouw is zeer bizar en wordt in veel illustraties en zelfs skeletopstellingen fout weergegeven. De romp is extreem afgeplat zodat hij de vorm heeft van een pillendoos. De bovenkant ervan heeft slechts een geringe en zeer geleidelijke kromming. De zijkanten vallen abrupt naar beneden weg. De poten zijn zeer kort en staan veel dichter op de middenlijn dan de uiterste zijkanten van de buikholte zodat die in vooraanzicht ver buiten de ledematen uitsteekt. De nek is tamelijk lang en dik en gaat naar voren over in een platte brede stompe kop waarvan de snuit sterk naar beneden gebogen is ten opzichte van de achterkant van de schedel. De staart heeft een dikke basis, maar versmalt al snel tot een hendel voor een hamervormige staartknots. Net als alle ankylosauriden, is Euoplocephalus zwaar bepantserd: vergroeide huidverbeningen vormen beenplaten die de staartbasis, romp, nek en kop volledig van boven bedekken. Op die platen staan rijen stekels.
Oudere diagnoses van Euoplocephalus zijn door de nieuwe inzichten achterhaald geworden. Arbour stelde in 2013 een nieuwe lijst kenmerken op waarin Euoplocephalus zich van verwante soorten onderscheidt. Deze eigenschappen zijn op zich niet uniek maar wel in combinatie. Euoplocephalus mist vergeleken met Anodontosaurus lambei en Scolosaurus cutleri ronde vastgegroeide huidverbeningen aan de bases van de hoorns op het squamosum en het quadratojugale. Hij mist vergeleken met Anodontosaurus lambei kleine pantserplaten op de voorste halsberg aan de bases van de grote osteodermen, maar heeft juist wel in bovenaanzicht halfronde grote osteodermen op de staartknots. Hij verschilt van Dyoplosaurus acutosquameus in het bezit van zijwaarts uitstekende sacrale ribben. Hij verschilt van Scolosaurus cutleri in het bezit van gekielde osteodermen met een ovale of ronde basis op de bovenkant en zijkanten van de eerste halsberg en in het bezit van een korter achterblad van het darmbeen. Euoplocephalus onderscheidt zich van Ankylosaurus magniventris in het bezit van voorwaarts gerichte neusgaten en door het ontbreken van een doorlopende kiel tussen de hoorn op het squamosum en de supraorbitalia.
Skelet
[bewerken | brontekst bewerken]Schedel
[bewerken | brontekst bewerken]
De schedel van Euoplocephalus is een gedrongen constructie, breder dan lang. Bij het grootste bekende exemplaar, specimen AMNH 5403, is het cranium 411 millimeter lang en 478 millimeter breed, gemeten over de quadratojugale hoorns. Het achterhoofd vormt in bovenaanzicht een bijna rechte rand waarvan de uiteinden, bekroond door hoorns op de squamosa, ver uit elkaar liggen. Het breedste punt van de bovenkant van de schedel ligt echter bij sommige exemplaren meer naar voren toe, boven de oogkassen die bovenop ook weer van hoorns voorzien zijn; de lijn tussen deze uitsteeksels en de hoorns op de squamosa is hol. Naar voren toe loopt de schedel uit in een korte stompe snuit. Dat levert in bovenaanzicht geen perfect tongvormig profiel op want de zijden ervan zijn wat ingevallen. De omtrek van de schedel als geheel is dus meer zeshoekig — maar is ook vergeleken met een afgeknotte gelijkzijdige driehoek. De voorkant van de snuit is relatief erg breed — ook vergeleken met veel verwante soorten — en in bovenaanzicht licht bol.

Van de buitenkant af is de indeling van de schedelelementen lastig waarneembaar doordat de kop wordt bedekt door een asymmetrisch mozaïek van beenplaatjes die per individu verschillen en tegenwoordig naar een suggestie van William Blows caputegulae genoemd worden, 'koptegeltjes'. Het gaat volgens de standaardinterpretatie om aan de schedel vastgegroeide huidverbeningen; de vergroeiing is zo hecht, de normale beennaden uitwissend, dat ook wel gesuggereerd is dat om een differentiatie van de schedeldelen zelf zou kunnen gaan, onder invloed van opliggende hoornschubben. De structuur hiervan is vooral complex en variabel boven de neusbeenderen; meer naar achter boven de voorhoofdsbeenderen en wandbeenderen versmelten de caputegulae tot een doorlopende beenplaat. Deze sluit het bovenste slaapvenster volledig. Het onderste slaapvenster is nog aanwezig maar geheel verborgen onder de hoorns van de achterste zijkant. Door de vergroeiingen zijn de precieze begrenzingen van het neusbeen, het traanbeen, het prefrontale, de vermoedelijk drie supraorbitalia, het postorbitale, het voorhoofdsbeen, het wandbeen en het jukbeen niet vaststelbaar. Alleen op de onderzijde van deze sectoren zijn soms nog beennaden aanwezig, maar deze is bij Euoplocephalus nooit volledig uitgeprepareerd. Op de bovenste achterhoek draagt het squamosum een hoorn van variabele lengte, waarvan de basis door diepe groeven gescheiden wordt van de plaat op het achterste schedeldak. Het squamosum is niet vergroeid met het onderliggende verticale quadratum, maar enige beweging tussen deze elementen lijkt niet mogelijk te zijn geweest. Het quadratum eindigt onderaan in de twee knobbels van het bovenste kaakgewricht. Op het quadratojugale bevindt zich een enorme platte driehoekige afhangende hoorn die naar beneden toe zijwaarts buigt.

In zijaanzicht is de schedel erg plat. Dat wordt wat verhuld door de grote afhangende hoorns op het quadratojugale, bij de onderste achterhoek van de schedel. De vrij kleine en hoekige oogkas bevindt zich laag onder een wat bollende sector van het schedeldak, gevormd door drie supraorbitalia. Bij verschillende exemplaren is in de bovenkant van de oogkas een kwartcirkelvormig los element gevonden, slechts zichtbaar door het verhemelte, dat een door de bepantsering naar binnen gedrongen rudiment kan zijn van het wenkbrauwbeen. Volgens Coombs functioneerde dit als een benig ooglid dat bij sluiting de oogkas, als zwak punt binnen de algehele bepantsering, afsloot. Vaak wordt het afgebeeld met ook nog een osteoderm erop, maar dit lijkt onverenigbaar met de functie van het vlezige ooglid. De bij dinosauriërs normale schedelopening vóór de oogkas, de fenestra antorbitalis, is geheel gesloten.

Schuin voor en onder de oogkas bevindt zich een kort bovenkaaksbeen dat een dus eveneens korte tandenrij draagt met kleine bladvormige tanden. De tandenrij is in bovenaanzicht sterk hol, met het diepst naar binnen gelegen punt op ongeveer een derde van de lengte van de voorste tand gelegen, en daarnaast als geheel ingesprongen ten opzichte van de zijkant van de schedel. De rij telt negentien tot vierentwintig tanden, gemiddeld tweeëntwintig, op een dunne beenrichel geplaatst. Het inspringen van de rij duidt erop dat een vlezige wang aanwezig was. Het waren niet alleen de tanden die het voedsel verknipten; het bovenkaaksbeen heeft aan de buitenzijde ook een opvallende snijrand. Aan de voorkant van het bovenkaaksbeen buigen het neusbeen en de praemaxilla bollend naar beneden toe zodat de snuitpunt extreem diep en schuin afhangt, bij sommige exemplaren wel een hoek van zestig graden met de tandenrij makend. De brede snuit draagt een schepvormige hoornsnavel, die achteraan in de snijranden van de bovenkaaksbeenderen uitloopt. De snavel moet hoog geweest zijn zoals blijkt uit het ontbreken van osteodermen op de tandeloze praemaxillae, de voorste snuitbeenderen. Deze laatste zijn van elkaar gescheiden door een opvallende diep ingesneden beennaad die aan de snuitrand een inkeping oplevert en meer naar binnen over korte stukken vaak niet gesloten is, zodat er openingen vallen. De snuit is binnenin gevuld door complexe luchtholten. De praemaxillae groeien aan hun onderste binnenkant uit in een in de lengterichting lopende interne richel die deze holten vooraan als een tussenschot scheidt. De aan iedere zijde dubbele neusgaten, dus met twee achter elkaar gelegen openingen, worden overhangen door de neusbeenderen. In vooraanzicht zijn de voorste neusopeningen breed en ovaal afgeplat, naar de buitenzijden schuin oplopend. De ovale achterste neusopeningen, van de voorste gescheiden door een verticaal schot afhangende van de achterste bovenste tak van de praemaxilla, zijn kleiner en juist naar buiten toe schuin naar beneden gericht. De voorste externe neusgaten leiden naar de normale neusholte, die vrij hoog is met een platte bovenkant, en waren kennelijk de belangrijkste luchtweg om adem te halen. De luchtweg gaat eerst schuin omhoog, de bolling van de snuit in, om daarna naar beneden en wat naar voren te buigen naar de interne neusgaten, de choanae. De achterste externe neusgaten leiden naar grote luchtholten boven de tandenrijen; twee luchtkamers zijn naast elkaar aanwezig. Tussen deze luchtholten en de neusholte bevindt zich aan iedere kant nog een luchtkamer met in vooraanzicht een driehoekige doorsnede. Volgens een studie uit 2011, waarbij specimen UALVP 47977 door middel van een CAT-scan onderzocht werd, deed de lucht bij het inademen ook nog zijwaarts deze kamer aan. Aan de achterkant van de neusholte bevindt zich achter een van het voorhoofdsbeen afhangend dwarsschot een aparte reukkamer waarvan de wanden sterk dooraderd waren. Hetzelfde onderzoek vond een raadselachtige dwarstunnel in het voorhoofdsbeen van specimen AMNH 5405, met een onbekende functie.

Het achterhoofd is plat met een brede en lage condylus occipitalis, de knobbel die contact maakt met de nek, onder een halfcirkelvormig achterhoofdsgat. De bovenrand wordt bekroond met een zware overhangende richel op de achterrand van de wandbeenderen. Onder de bovenrand bevindt zich een enkelvoudig supraoccitipale met middenin in een richel in de vorm van een omgekeerde 'Y' waarvan de onderste takken het achterhoofdsgat omsluiten. De zijuitsteeksels, de processus paroccipitales, zijn horizontaal en bijna perfect zijwaarts gericht. De uiteinden ervan raken de bovenzijden van de quadrata niet. De onderliggende delen van het achterhoofd en de hersenpan zijn sterk vergroeid zodat een stevig geheel ontstaat waarin de verschillende elementen nauwelijks meer te onderscheiden zijn. Onderaan de hersenpan hangen twee tubercula basilaria, in de vorm van lage richels. Daarvoor hangen twee stevige processus basipterygoidei. De voorkant van de hersenpan, het parasfenoïde, maakt contact met het achterste neustussenschot, volgens de studie uit 2011 via verbeende ethmoïde elementen, kraakbeenlagen die normaal het middelste van de schedel wat structuur geven. Bij geen enkel exemplaar zijn de stapes gevonden, de staafvormige verbindingen tussen het binnenoor en het trommelvlies, die trouwens bij alle Ankylosauria onbekend zijn. Van de hersenen zelf is bij specimen AMNH 5337 een natuurlijk afgietsel bewaard gebleven. Dit toont dat het brein horizontaal ligt, en compact is zonder een erg duidelijke onderverdeling, met vooraan korte stevige reuklobben. De kanalen voor de hersenzenuwen liggen ook vrij dicht op elkaar.
De structuur van de onderkant van de schedel is, in tegenstelling tot die van de bovenkant, goed waarneembaar en heeft daarom altijd sterk de aandacht getrokken van de onderzoekers. De binnenvleugels van de bovenkaaksbeenderen zijn met interne platen van de praemaxillae en de ploegschaarbeenderen versmolten tot een doorlopend secundair verhemelte. Dit loopt helemaal naar achteren door tot recht vóór de grote interne neusgaten, die de neusholten verbinden met de keelholte. De ploegschaarbeenderen vormen een onderste kiel op de middenlijn en scheiden beide interne neusgaten, zo een verbinding vormend met de achter deze openingen gelegen verhemeltebeenderen, die dan als het ware een tweede, achterste, deel van het secundaire verhemelte vormen. Deze vormgeving bij de ankylosaurinen wijkt sterk af van de situatie bij andere dinosauriërs die of helemaal geen benig secundair verhemelte hebben of een dat tot de snuit beperkt is. Het verhemeltebeen zelf heeft in het midden een kleine ronde opening die toegang geeft tot een naar achteren doorlopende holte, een typisch kenmerk van de ankylosauriden. Achter het verhemeltebeen bevindt zich een complex driestralig pterygoïde. Het brede hoofdlichaam ervan is verticaal georiënteerd en loopt naar de richel van de ploegschaarbeenderen. Achter hun raakpunt bevindt op de middenlijn een enkelvoudige driehoekige deuk tussen in de lengterichting lopende spleten op de gepaarde pterygoïden. Een ruwe zijtak loopt schuin naar voren, uitlopend in een klein ectopterygoíde naar de schedelrand, een andere korte brede tak schuin naar achteren naar het quadratum en is daarmee verbonden in een haaklas, dus met een opliggende beenplaat. Het pterygoïde is niet verbonden met de hersenpan; bij het levende dier lag er vermoedelijk kraakbeen tussen.
Onderkaken
[bewerken | brontekst bewerken]
De onderkaak, die maar van enkele exemplaren bekend is, is een krachtige en hoge maar ook zeer complexe structuur. Zij heeft een lengte van ongeveer zeventig tot tachtig procent van de schedel. In het midden ervan vormt het tanddragend bot, het dentarium of os dentale, een hoge kromming met vooral een steile voorzijde. Hierdoor kan de onderkaak aansluiten bij de gekromde onderkant van de schedel en diens afhangende snuit. De onderkaak moet echter ook een ondersnavel dragen die onder de bovensnavel van de snuit aansluit. Daartoe kromt aan de voorste basis van de bult van het dentarium een zeer dunne voorste tak weer naar boven toe. Deze tak draagt een in zijaanzicht zeer kort predentarium, de beenkern van de ondersnavel. Die kern wordt gedeeld met de andere onderkaak. Beide onderkaken zijn aldus verbonden door een zeer breed enkelvoudig schepvormig bot dat bedekt moet zijn geweest met een hoge hoornige schacht die in de bovensnavel paste. Het predentarium wordt meestal niet voorgesteld als een symfyse, samengroeiing, van beide onderkaakpunten, maar een dergelijke oorsprong wordt gesuggereerd door een uitsteeksel in het midden van het element. De raakpunten tussen het predentarium en de voorste takken van de dentaria zijn overdwars diep ingesneden. Gecombineerd met de lage hoogte van die takken is dat gezien als een aanwijzing dat de verbinding zeer losjes was en een verticale beweging van de ondersnavel toestond.
Ondanks de lange kromme van het dentarium, draagt zijn maar eenentwintig tanden, dus gemiddeld minder dan het bovenkaaksbeen. Omdat de tandenrij van het bovenkaaksbeen in bovenaanzicht sterk hol is, moet ook het dentarium in het midden naar binnen plooien. Een horizontale rij grote aderkanalen loopt over de buitenste wand van het dentarium. Aan de binnenzijde is een rij kleine putjes te zien die de posities van de vervangingstanden aangeven. Daaronder loopt een diepe groeve, de fossa Meckeliana, die overspannen wordt door het spleniale, de lange beenband aan de binnenzijde van de onderkaak. De onderkant van de buitenzijde wordt gevormd door een angulare, dat diep naar beneden uitsteekt en zo ruw is dat het vermoedelijk direct verbonden was met huidverbeningen, maar zich niet om de onderkant van de kaak heen wikkelt. De achterste buitenste zijwand wordt gevormd door een groot driehoekig surangulare. Dit wordt doorboord door een van voren naar binnen lopend foramen maar de onderkaak heeft geen groot zijvenster. Aan de binnenzijde vormt het surangulare volgens Vickaryous de achterkant van het lage dunne hefboomsuitsteeksel, de processus coronoides, waarvan de voorkant gevormd zou worden door een apart os coronoides; volgens Coombs is dit echter afwezig. Bij specimen UALVP 31 is de processus coronoides opvallend hoger dan bij alle andere exemplaren, ook als men rekening houdt met compressie. Onder het surangulare en boven het spleniale ligt aan de binnenzijde een lang prearticulare dat ver naar achteren doorloopt en aan de achterzijde een retroarticulair uitsteeksel vormt. Dit is hoger dan lang, overdwars afgeplat en aan de achterkant verruwd. Het prearticulare vormt ook de achterste binnenwand van de opening waardoorheen de sluitspieren de holle binnenkant van de onderkaak ingaan. Achter die opening steekt het articulare omhoog, het onderste kaakgewricht vormend. Het gewrichtsvlak is overdwars breed en is door een lage richel in een voorste en achterste uitholling verdeeld. Volgens een studie van Vickaryous uit 2001 verliep de sluiting van de onderkaken in twee fasen: een verticale beweging gevolgd door een achterwaartse beweging. Deze laatste zou ook de ondersnavel ten opzichte van de bovensnavel naar achteren hebben laten bewegen en zou de knobbels van het quadratum in de achterste gewrichtsholte getrokken hebben.
Tanden
[bewerken | brontekst bewerken]De tanden van Euoplocephalus zijn opvallend klein ten opzichte van de schedel, met een maximale lengte en breedte van slechts 7,5 millimeter. De premaxillaire tanden ontbreken geheel. Gezien de variabiliteit in het aantal maxillaire tanden van de bovenkaak varieert het totaal voor de kop tussen de tachtig en negentig. Tussen de maxillaire en dentaire tanden is geen verschil in vorm. De tandbasis is wel wat verbreed maar een echt cingulum, opstaande rand, ontbreekt. De tandkroon heeft zeventien vertandingen. De vertandingen zijn aan de snijrand plat. Ze zijn van andere vertandingen gescheiden door diepe inkepingen die uitlopen in verticale groeven, zowel aan de buitenzijde als de binnenzijde van de tand. Naar beneden lopen die groeven eerst naar elkaar toe en dan weer uiteen. De versmalling in het midden komt overeen met het wijdere gedeelte van de vertanding die een positie lager op de snijrand staat. Alle vertandingen hebben zulke versmallingen die een typisch kenmerk vormen van Euoplocephalus en zijn Noord-Amerikaanse verwanten uit het Campanien. De tand waarop Palaeoscincus asper gebaseerd is, werd vanwege een hiermee overeenkomende morfologie aan Euoplocephalus tutus gelijkgesteld. Een middelste en grootste vertanding vormt het spits van de tandkroon.
Postcrania
[bewerken | brontekst bewerken]
Wervelkolom
[bewerken | brontekst bewerken]De indeling van de wervelkolom is niet met zekerheid bekend. Er zijn minstens zeven halswervels en elf niet-vergroeide ruggenwervels. De laatste vier ruggenwervels zijn meestal vergroeid met elkaar en het heiligbeen en zijn dus dorsosacrale wervels. Het aantal sacrale wervels in engere zin beloopt drie. Achter het heiligbeen is altijd nog minstens één caudosacrale wervel van de staart vergroeid zodat het totaal aan sacrale wervels in ruime zin meestal acht bedraagt. Bij sommige exemplaren ligt dit echter hoger; vermoedelijk oudere dieren. Specimen ROM 1930 heeft drie caudosacrale wervels en wellicht nog een extra dorsosacrale wervel; specimen TMP 1982.9.3 heeft vier caudosacrale wervels. Het aantal staartwervels ligt vermoedelijk rond de dertig waarvan er een tiental vergroeid zijn tot de hendel van de staartknots.
Van de halswervels is de atlas onbekend, hoewel hij er als voorste wervel natuurlijk wel aanwezig was. De tweede wervel, de draaier, is de langste onder de halswervels. Hij was, gezien een massief naar voren uitstekend odontoïde met een ondiepe in doorsnede U-vormige groeve op de bovenkant, stevig met de atlas verbonden. De draaier heeft nekribben, een V-vormig naar achteren gericht doornuitsteeksel, en overhangende achterste gewrichtsuitsteeksels met ovale en van voor naar achteren lange facetten. De overige halswervels zijn even breed, of iets breder, als lang met aan beide zijden ronde en holle vlakken op de dus amficoele wervellichamen. Hun doornuitsteeksels zijn aan de achterzijde diep uitgehold zodat ze in bovenaanzicht een U-vorm hebben. De takken daarvan gaan naar boven toe verder uiteen staan zodat het uitsteeksel in vooraanzicht de vorm heeft van een op zijn punt staande driehoek. De voorste gewrichtsuitsteeksels overhang de voorrand van het wervelcentrum en staan wijd uiteen, verbonden door een lage overdwarse beenschort met een horizontale bovenkant. Het ruggenmergkanaal heeft een vierkante doorsnede. De achterste gewrichtsuitsteeksels overhangen niet en zijn ook niet verbonden met enige epipofysen. Het zijuitsteeksel staat laag op de wervelboog en is naar beneden en zijwaarts gericht. Het onderste ribgewricht, de parapofyse, ligt vooraan op de zijkant van het wervellichaam.
Bij de ruggenwervels zijn de doornuitsteeksels simpele bladen, naar achteren gericht en in zijaanzicht rechthoekig en dun. De ruwe uiteinden zijn iets overdwars gezwollen. De wervellichamen zijn spoelvormig en hebben ronde uiteinden. Het ruggenmergkanaal is hoog en ovaal in doorsnede. De zijuitsteeksels zijn zowel lang als breed. Ze dragen een diapofyse, bovenste ribgewricht, in de vorm van een omgekeerde driehoek op het uiteinde. De parapofyse is rond of ovaal; nog functionerend als een gewricht in de voorste ruggenwervels, wordt het tot een beennaad in de achterste, een teken dat daar de ribben vastgroeien als onderdeel van de algehele onbeweeglijkheid die het achterste romp kenmerkt. De voorste gewrichtsuitsteeksels zijn sterk omhoog gericht en staan nauw opeen, gescheiden door een U-vormige trog. De achterste gewrichtsuitsteeksels zijn versmolten tot een enkel penvormig uitsteeksel op de middenlijn, opnieuw een aanwijzing voor verstarring.
De caudosacrale wervels zijn nier altijd met hun wervellichamen met het heiligbeen vergroeid, maar bij alle sacrale wervels zijn de doornuitsteeksels versmolten tot een doorlopende supraneurale plaat waarin ook de gewrichtsuitsteeksels zijn opgenomen. Aan de golving in de soms wat verbrede bovenrand van die plaat zijn de posities van de wervels nog te herkennen. De groeve op de onderkant van het heiligbeen is ondiep of afwezig. De drie sacrale wervels in engere zin hebben in doorsnede meer rechthoekige en dikkere zijuitsteeksels die geen T-vormige dwarsdoorsnede hebben zoals bij de dorsosacrale wervels.
Bij de staartwervels hebben de wervellichamen typisch een ronde of ovale doorsnede. De doornuitsteeksels en chevrons zijn plaatvormig en verengen naar hun uiteinde toe. De doornuitsteeksels zijn matig lang. Bij de middenstaart zijn ze meer naar achteren gericht en de chevrons worden daar sledevormig. Vanaf ongeveer de twaalfde vrije staartwervel vormen de chevrons een doorlopende onderkant. De doornuitsteeksels verdwijnen en de lage bovenkant van de wervel wordt een stevige, lange, en naar achteren stekende punt die tussen twee naar voren stekende punten van de achterliggende wervel past. Zo wordt het dunne staartuiteinde verstijfd zonder alle flexibiliteit te verliezen. Eenzelfde functie hebben bundels lange verbeende pezen die naast de wervellichamen lopen. De staart eindigt in een zware staartknots gevormd door grote osteodermen.
Ledematen
[bewerken | brontekst bewerken]Het schouderblad is breed en peddelvormig met een iets verwijd uiteinde. De achterrand is licht hol. Op de onderste voorrand bevindt zich een spits uitsteeksel, de processus acromialis, dat het hoofdlichaam zijwaarts overhangt en aan de onderzijde een driehoekige groeve heeft, waarlangs het in grootte afneemt. Er is een diep aanhechtingspunt voor de Musculus triceps longus caudalis. Aan de binnenzijde is er een flink opstaande richel op de grens met het ravenbeksbeen. Aan de achterste buitenzijde, waar schouderblad en ravenbeksbeen samen gelijkelijk aan het schoudergewricht bijdragen, heeft het schouderblad een driehoekig naar beneden gericht uitsteeksel op de onderrand. Het ravenbeksbeen heeft een vierkant profiel met een rechte achterrand en een haakvormig uitsteeksel richting borstbeen. Het wordt doorboord door een rond foramen.
Het opperarmbeen is robuust en zandlopervormig door verbrede uiteinden en een smalle schacht. Dit duidt op een zeer zware musculatuur. Een enorme deltopectorale kam beslaat aan de voorste binnenkant de bovenste tweeënveertig graden van de schachtlengte. Hij gaat onderaan niet geleidelijk in de schacht over, maar buigt daar verder naar voren toe, een dikke knop vormend. De kam is diep gegroefd voor aanhechting van de Musculus supracoracoideus en de Musculus pectoralis major. De gewrichtskop van het opperarmbeen is rond in bovenaanzicht, iets driehoekiger van de binnenkant bekeken. Tussen kop en kam ligt vooraan een brede ondiepe trog, de fossa bicipitalis. Aan de bovenste binnenzijde van de schacht ligt een dikke bult. Achter de kop is de achterrand glad. De onderste gewrichtsknobbels zijn groot en sterk overdwars verbreed; de buitenste is wat forser dan de binnenste. De inkeping ertussen is ondiep en afgerond. Bovenaan loopt deze uit in een achterste groeve, de fossa olecrani die ondiep is en driehoekig. alle uiteinden van het opperarmbeen tonen diepe oppervlaktegroeven die wijzen op dikke kappen van kraakbeen. Het spaakbeen is robuust met een wijd uitstaand hol bovenste gewrichtsvlak en een onderste uiteinde dat in vooraanzicht een ruwe stompe punt vormt. De ellepijp heeft aan de achterste bovenkant een krachtig ruw uitsteeksel, de processus olecrani. Verder is dit element slecht bekend evenals de pols en hand.

Het bekken is breed en lang met naar voren toe wijd uiteen waaierende darmbeenderen om de enorm brede buikholte te ondersteunen. Het voorblad van het darmbeen loopt daartoe helemaal door tot het breedste punt van de buik. Dit is ook de reden voor het grote aantal dorsosacrale wervels: de achterste rugwervels worden 'ingevangen' om met hun zijuitsteeksels weer dat voorblad te ondersteunen. In zijaanzicht is het darmbeen matig hoog; in bovenaanzicht vormt het een brede, naar voren versmallende, plaat, die naar bezijden toe naar beneden omkrult. Het achterblad is bij ankylosauriden erg kort doordat het heupgewricht sterk naar achteren is gelegen teneinde de achterpoot de vrijheid te verschaffen om ondanks de brede buikholte naar voren te bewegen. Bij Euoplocephalus is het achterblad iets langer dan de doorsnede van het heupgewricht zelf. Het schaambeen is onbekend. Het zitbeen is een kort, bijna verticaal naar beneden krommend element. Aan de bovenkant is het breed, zonder een duidelijke splitsing in een tak richting darmbeen en een tak richting schaambeen. Aan de buitenzijde ligt een trog en die gaat naar boven over in een beenplaat die bijdraagt aan de binnenwand van het heupgewricht dat helemaal aan de binnenzijde gesloten is en niet geperforeerd. Het hoofdlichaam gaat naar onderen tie abrupt in de schacht over, die overdwars is afgeplat en in achteraanzicht een golvend profiel heeft. De voorrand en achterrand lopen over hun volle lengte vrijwel parallel en de onderkant is recht zodat de schacht bandvormig is in zijaanzicht.
Het dijbeen is kort maar stevig met een rechte schacht en een ovale dwarsdoorsnede. De kop is rond en gaat zonder nek over in de even hoge bovenkant van de schacht, de trochanter major. Aan de achterkant van de schacht bevindt zich iets onder het midden een lage verruwing die overeenkomt met de vierde trochanter, de aanhechting van de retractorspier van de staart, de Musculus caudofemoralis longus. De lage positie duidt erop dat achterpoot gebouwd is voor kracht, niet snelheid. De onderste gewrichtsknobbels zijn naar achteren verbreed; de binnenste is iets groter dan de buitenste. Er is een ondiepe achterste groeve tussen de knobbels. Het scheenbeen is korter dan het dijbeen en heeft sterk verbrede uiteinden. Onderaan is het bot in vooraanzicht maar iets smaller dan bovenaan, maar in zijaanzicht twee maal zo nauw. Dit duidt op een brede voet die echter zelf slecht bekend is; vroeger werd het juveniele exemplaar AMNH 5266 hiervoor als informatiebron gebruikt, maar Arbour wees dit aan Anodontosaurus toe. De voet heeft drie dragende tenen en de voetklauwen zijn hoefvormig met een U-vormig in plaats van driehoekig profiel in bovenaanzicht.
Osteodermen
[bewerken | brontekst bewerken]
Caputegulae op de schedel
[bewerken | brontekst bewerken]Euoplocephalus is gepantserd met huidverbeningen ofwel osteodermen, die geen deel uitmaken van het eigenlijke skelet. De osteodermen van de schedel, de caputegulae, zijn het best bekend omdat ze, volledig met de schedelelementen vergroeid, zich meestal nog in hun oorspronkelijke positie bevinden. Hun structuur werd echter door eerdere auteurs verwaarloosd omdat die eraan wanhoopten in het chaotische geheel enige ordening te kunnen aanbrengen. Tegenwoordig zijn echter jonge ankylosauriden uit Azië bekend die laten zien dat de kop op jonge leeftijd bedekt was met grotere platen die eenduidig in verband te brengen zijn met bepaalde schedelbotten. De kleinere plaatjes die op latere leeftijd ontstaan, hebben hierin hun herkomst en kunnen aan de hand hiervan worden ingedeeld. Arbours beperking van het materiaal van Euoplocephalus leidde daarbij tot een grotere eenvormigheid zodat ze in 2013 in haar revisie van het geslacht een poging waagde tot een gedetailleerde beschrijving.
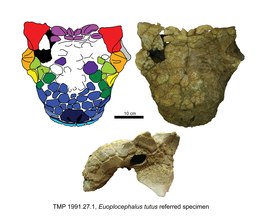
Boven de vermoedelijke positie van de op zichzelf niet bewaard gebleven bovensnavel liggen de voorste plaatjes van het neusbeen. Die vormen ook de rand van de externe neusgaten en zijn ruwer dan meer naar achteren gelegen caputegulae. De achterste plaatjes van het neusbeen zijn gepaard en min of meer rechthoekig van vorm. Op de achterrand van de zone van de neusbeenderen — door het ontbreken van een beennaad met de voorhoofdsbeenderen gaan de verschillende sectoren vloeiend in elkaar over — ligt een grote zeshoekige plaat op de middenlijn. Nog verder naar achteren wordt deze zone en die van de voorhoofdsbeenderen gekenmerkt door een mozaïek van kleine ronde, rechthoekige of zeshoekige plaatjes die per individu verschillen. Aan de achterste zijkant van het neusbeen vormt een grote gekielde osteoderm de bovenhoek van de achterste snuit. Erachter ligt nog zo'n hoekplaat, boven het traanbeen, maar die loopt minder ver op het schedeldak door. De bovenste rand van de oogkas wordt bekroond door twee grotere in bovenaanzicht driehoekige supraorbitale osteodermen, een voorste en een achterste, die zijwaarts uitstekende in elkaar verlengde liggende kielen hebben die een doorlopende rand vormen met eenzelfde oriëntatie als de hoorn op het squamosum, dus schuin naar achteren en bezijden gericht. De gedeelde richting kan een esthetische of defensieve functie hebben gehad. De achterste hoek ervan is bij twee exemplaren, specimina TMP 1991.127.1 en UALVP 31, het breedste punt op de schedel. Desalniettemin is de achterste supraorbitale osteoderm bij deze fossielen meer afgerond en lager; hij heeft ook een opvallende dwarsgroeve. Het geheel van de bovenste oogrand, driehoekig in zijaanzicht, steekt bij deze specimina opvallend verder naar achteren uit. Rond de rest van de oogkas is bij sommige exemplaren een rand van kleine osteodermen aanwezig. Achter de postorbitalia, in de zone van de wandbeenderen, zijn geen duidelijke individuele plaatjes meer te onderscheiden in wat een enkele versmolten beenplaat is geworden. Op de achterrand van de wandbeenderen echter ligt een dwarskam die de overgang vormt met het achterhoofd en deze is bezet met vier vierkante en of rechthoekige caputegulae, in twee paren waarvan het middelste paar het kleinste is. Deze osteodermen beslaan meestal niet de volle randlengte. Vaak zijn beide zijden wat bollend met in het midden een inkeping.

De hoorn op het squamosum is tussen de verschillende exemplaren sterk verschillend in lengte, scherpte en oriëntering. Meestal is hij driehoekig in zowel bovenaanzicht als zijaanzicht. In sommige exemplaren is de achterkant ervan een voortzetting van de dwarskam op het bovenste achterhoofd; bij andere liggen deze hoorns vóór of steken ver achter de achterrand. De hoorn is altijd enigszins naar achteren en zijwaarts gericht; de oriëntatie verschilt vooral in het verticale vlak: de punt kan schuin naar beneden maar ook schuin naar boven gericht zijn. Meestal is bij hetzelfde individu de vorm en positie links en rechts identiek. Ook de grotere hoorn op het quadratojugale is sterk variabel. De punt ervan kan vrij scherp zijn of juist stomp afgerond. De hoorn kan recht naar beneden gericht zijn of schuin naar achteren. Bij sommige exemplaren bevinden zich aan de bases van de squamosale of quadratojugale hoorns kleine ronde beenplaatjes.
Pantserplaten van de romp
[bewerken | brontekst bewerken]De bepantsering van de romp is in 1982 diepgaand beschreven door Kenneth Carpenter. Deze baseerde zich hierbij echter op het holotype van Scolosaurus dat toen nog als een exemplaar van Euoplocephalus werd gezien. Arbour gaf in 2013 een herziene beschrijving. De halsbergen op de nek hebben een complexe structuur. Bovenop bevindt zich een laag van normale osteodermen, maar dieper ligt een dikke beenband die het geheel stevigheid en sterkte gaf. Het bestaan van zo'n band is een problematisch gegeven omdat het niet duidelijk is of het om een huidverbening gaat, een verbening van kraakbeenlagen of misschien zelfs om een nieuwvorming van het skelet. Dat het wellicht niet om een huidverbening gaat, kan blijken uit het feit dat het bot aan het oppervlak niet ruw is maar glad en intern een geweven in plaats van een sponsachtige structuur heeft. De band is onderverdeeld in verschillende rechthoekige hol gebogen platen die hecht verbonden zijn met gekartelde beennaden. Typisch zijn er zes per ring, in drie paren. Meestal ligt er op de plaat een grote centrale osteoderm; soms zijn er geen of twee grote osteodermen aanwezig. De osteoderm kan wel, gedeeltelijk of geheel niet vastgegroeid zijn met de beenband. Deze kenmerken worden door de meeste ankylosauriden gedeeld; bij Euoplocephalus draagt het centrale paar een grote ovale osteoderm met in de lengterichting een hoge kiel, terwijl het paar dat de zijhoeken vormt een kleinere osteoderm heeft met een gegolfde kiel. De platen aan de onderste zijkant zijn soms glad, soms is er een gladde afgeplatte osteoderm aanwezig. Volgens Arbour behoren exemplaren met ronde osteodermen aan de bases van de grote osteodermen of met kielen die zo groot zijn dat ze naar achteren toe de achterrand van de halsberg overhangen, niet tot het materiaal van Euoplocephalus. De vorm van de voorste halsberg lijkt op die van de achterste, maar de dimensies zijn kleiner.

De indeling van het romppantser is niet goed bekend. Vermoedelijk is er een heupschild boven het bekken en daarvóór liggen overdwars georiënteerde banden over de borstkas. Deze banden bestaan, anders dan de halsbergen, niet uit een diepe beenlaag, maar zijn opgebouwd uit samengegroeide beenschubben. Rijen gekielde osteodermen lopen over de romp naar achteren. Zulke rijen zijn er ook over de staartbasis die eveneens pantserbanden heeft. Specimen ROM 1930 heeft het staartpantser bewaard waarop ovale osteodermen aanwezig zijn met een lengte van vijftien centimeter. Het exemplaar omvat daarnaast honderden kleine beenschubben met een diameter van minder dan vijf millimeter. Specimen TMP 1997.132.01 heeft bij het opperarmbeen grote ronde gekielde osteodermen bewaard met een doorsnede van twintig centimeter. Op de onderarm staan kleinere osteodermen die een langgerektere basis hebben en stekelvormige kielen die de achterrand overhangen.
Staartknots
[bewerken | brontekst bewerken]De osteodermen van de staartknots maken het grootste deel van de massa van dat orgaan uit. Links en rechts bevindt zich een grote afgeplatte lob die bij Euoplocephalus in bovenaanzicht een rond profiel heeft, niet een driehoekig zoals bij sommige verwanten. De lob is even lang als breed of langer dan breed. Losse driehoekige staartknotsen die breder zijn dan lang acht Arbour niet tot het zekere materiaal van Euoplocephalus te behoren. De grote lob kan in zijaanzicht vrijwel bolvormig zijn maar ook meer lensvormig; dit is hiervan afhankelijk of de kiel zich op de rand of de bovenkant vormt. Het achterste uiteinde van de staart wordt gevormd door een paar kleinere osteodermen; meestal zijn die gepaard met een centrale osteoderm bovenop, maar de structuur varieert per individu en is door beschadigingen vaak niet goed waarneembaar.
Oudere illustraties tonen soms stekels op de staartknots. Dit is een gevolg van een oude reconstructie door Franz Nopcsa die, zich niet bewust van de speciale structuur van de hendel, stekelvormige osteodermen op de staart afbeeldde die tot de knots doorliepen. Later illustratoren zetten toen die stekels ook op de knots zelf.
Fylogenie
[bewerken | brontekst bewerken]In 1910 bracht Lambe Euoplocephalus onder bij de Stegosauria, maar dat begrip omvatte toen alle bepantserde vormen en was dus veel ruimer dan wat er nu onder verstaan wordt. In 1917 volgde een plaatsing door Charles Whitney Gilmore in de Ankylosauridae, die nog steeds wordt aangenomen. Volgens een cladistische analyse van Arbour uit 2013 heeft Euoplocephalus een vrij basale positie, onder in de stamboom van de Ankylosauridae, misschien met Anodontosaurus een klade, aftakking, vormend die het zustertaxon is van Ankylosaurus. De uitkomst hiervan bleek echter zwak ondersteund en was dus zeer onzeker; als slechts enkele kenmerken in een andere richting hadden gewezen, was de gevonden stamboom geheel anders uitgevallen.
De stamboom van de Ankylosauridae volgens een eerdere analyse van Susannah Maidment uit 2011 toont het volgende kladogram.
| Ankylosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Levenswijze
[bewerken | brontekst bewerken]Euoplocephalus leefde in een vrij natte kustvlakte op de oostkust van Laramidia met warme zomers maar koele winters. Het kustgebied werd gekenmerkt door moerassen en brede rivieren, meer landinwaarts waren er dicht beboste drogere heuvels. Uit de Dinosaur Park Formation is een groot aantal dinosauriërs bekend.
Volgens Arbour is Euoplocephalus voornamelijk uit de onderste en wellicht uit de vroege middelste Dinosaur Park Formation bekend en zijn latere ankylosauride vondsten van Anodontosaurus, zoals getoond wordt in het volgende diagram. De tijdsaanduidingen daarin zijn bij benadering omdat het vaak niet mogelijk is vindplaatsen precies te lokaliseren of dateren.

Het direct opeenvolgend voorkomen in hetzelfde gebied van eerst Scolosaurus, dan Euoplocephalus en ten slotte Anodontosaurus, vormen die in bouw zeer gelijkend zijn, suggereert sterk dat het hier om een doorlopende populatie of een chronospecies gaat, maar het zou ook achtereenvolgende afsplitsingen kunnen betreffen. Omdat een directe afstamming door een gebrek aan fossielen en een onvoldoende nauwkeurige datering niet bewezen kan worden, gaat men er in de moderne fylogenetische methodiek bij grote gewervelde dieren steeds van uit dat het aparte takken zijn.

Euoplocephalus zocht zijn eten vermoedelijk dicht bij de grond. Zijn lange nek stond nog een verticaal bereik van bijna twee meter toe, maar zich op zijn achterpoten verheffen kon hij niet. De brede bek duidt erop dat hij zoveel mogelijk laagwaardig plantenmateriaal met zijn snavel afbeet om dat, wellicht met behulp van gastrolieten ofwel maagstenen, in de enorme buikholte te verteren. Door een zeer lang darmtraject konden de darmbacteriën ook uit moeilijk te verwerken planten toch nog redelijk wat energie halen. Omdat hij bijna alles wat groeide kon eten, hoefde Euoplocephalus niet ver te lopen. Zijn lichaamsbouw wijst niet op een zeer actief leven. In zijn leefgebied kwam ook de nodosauride Edmontonia voor. Een nicheverdeling zorgde ervoor dat ze elkaar niet te veel beconcurreerden: Edmontonia had een smallere bek, at vermoedelijk hoogwaardiger kost, en was, getuige zijn veel langere poten, ook veel actiever. Dat blijkt zelfs uit hun wijze van vechten: de nodosauride ramde aanvallers met de lange stekels op zijn voorste romp. In hun habitat leefde de grote tyrannosauride Gorgosaurus. Euoplocephalus verdedigde zich daartegen juist passiever door zich met zijn korte poten schrap te zetten tegen de grond. De zeer zwaar gespierde armen bewogen de voorste romp heen en weer, wat de staart in tegenovergestelde richting deed zwiepen. Een enkele inslag van de hamervormige staartknots was voldoende om poten, borstkas of kop van een roofsauriër te verbrijzelen. Het verstijfde staartuiteinde verminderde de snelheid wel iets, maar vergrootte de kans op een zware schade toebrengende plastische vervorming in plaats van elastische vervorming van de weefsels van het ongelukkige slachtoffer. De staart kon slechts de achterkant van het lichaam dekken en dat verklaart waarom de voorste romp, de nek en de kop zo zwaar gepantserd waren. De gekielde osteodermen werkten misschien voornamelijk afschrikwekkend, maar de scherpe hoornlaag erop kon ook de huid van de aanvaller opensnijden of zijn tanden doen breken of afketsen.

Coombs had in 1978 overigens een veel optimistischer kijk op de beweeglijkheid van Euoplocephalus en meende dat het dier net zo snel had kunnen rennen als een neushoorn of een nijlpaard. Deze hoefdieren kunnen echter galopperen, iets waartoe Euoplocephalus naar huidige inzichten met zijn stijve romp en korte pootjes niet in staat was. Een draf van zo'n vijftien kilometer per uur moet echter tot de mogelijkheden hebben behoord.
Een rol in de verdediging werd wellicht ook gespeeld door het ingewikkelde ademhalingstraject, dat wel gezien is als een compensatie voor het verkorten van de snuit, teneinde de oppervlakte van het reukepitheel te handhaven. Kennelijk bleef de reuk een essentieel zintuig. Voor een voedselgeneralist als Euoplocephalus, die vermoedelijk alles vrat wat voor zijn neus opdook, zou ruiken echter minder belangrijk moeten zijn geweest bij het opsporen van voedsel. Het ware belang zou dus gelegen kunnen hebben in het op tijd waarnemen van de geur van een aanvaller. De dichte begroeiing maakte de reuk daarvoor een beter instrument dan het zicht.
Van Euoplocephalus zijn geen eieren, nesten, broedkolonies of seksuele dimorfie bekend. Het is niet duidelijk of het dier solitair leefde of in kudden. Het feit dat de fossielen nooit in grote opeenhopingen zijn gevonden, is wel gezien als een aanwijzing voor het eerste. Een alternatieve verklaring van de lange luchtwegen, dat ze dienden om een geluidssignaal voort te brengen, zou echter kunnen wijzen op een zekere mate van sociaal gedrag. Eveneens is het mogelijk dat de bewapening mede voor gevechten binnen de soort zelf gebruikt werd. Ook jongen zijn niet aanwezig onder het materiaal dat Arbour erkent. Wel wees zij erop dat de grootste exemplaren wat afgesleten hoorns en ruwere osteodermen hebben. Dat zou een gevolg kunnen zijn van slijtage of absorptie van deze structuren op latere leeftijd.
Literatuur
- L.M. Lambe, 1902, "New genera and species from the Belly River Series (mid-Cretaceous)", Geological Survey of Canada Contributions to Canadian Palaeontology 3(2): 25-81
- L.M. Lambe, 1910, "Note on the parietal crest of Centrosaurus apertus and a proposed new generic name for Stereocephalus tutus", The Ottawa Naturalist 24: 149-151
- Hennig, E., 1915, Stegosauria: Fossilium Catalogus I, Animalia pars 9, 16 pp
- Kuhn, O., 1964, Ornithischia: Fossilium Catalogus, I: Animalia, Pars 105, 80 pp
- Coombs, W.P. Jr., 1971, The Ankylosauria, Dissertatie Columbia University, New York, 487 pp
- Coombs, W.P. Jr., 1972, "The bony eyelid of Euoplocephalus (Reptilia, Ornithischia)", Journal of Paleontology, 46: 637–650
- Coombs, W.P. Jr., 1978, "An endocranial cast of Euoplocephalus (Reptilia, Ornithischia)", Palaeontografische Abteilung A 161: 176–182
- Coombs, W.P. Jr., 1978, "The families of the ornithischian dinosaur order Ankylosauria", Palaeontology 21: 143–170
- Carpenter, K. 1982, "Skeletal and dermal armor reconstruction of Euoplocephalus tutus (Ornithischia: Ankylosauria) from the Late Cretaceous Oldman Formation of Alberta", Canadian Journal of Earth Sciences 19: 689–697
- Rybczynski, N., Vickaryous, M.K. 2001, "Evidence of complex jaw movement in the Late Cretaceous ankylosaurid Euoplocephalus tutus (Dinosauria: Thyreophora)" pp 299–317 in: Carpenter K, editor. The armored dinosaurs, Bloomington: Indiana University Press
- Penkalski, P., 2001, "Variation in specimens referred to Euoplocephalus tutus" pp 363–385 in: K. Carpenter, editor. The armored dinosaurs, Bloomington: Indiana University Press
- Vickaryous, M.K. & Russell, A.P. (January 2003). A redescription of the skull of Euoplocephalus tutus (Archosauria: Ornithischia): a foundation for comparative and systematic studies of ankylosaurian dinosaurs. Zoological Journal of the Linnean Society 137 (1): 157–186. DOI: 10.1046/j.1096-3642.2003.00045.x.
- Miyashita, T., Arbour, V.M., Witmer, L.M., Currie, P.J., 2011, "The internal cranial morphology of an armoured dinosaur Euoplocephalus corroborated by X-ray computed tomographic reconstruction", Journal of Anatomy 219: 661–675
- Arbour V.M. and Currie P.J., 2013, "Euoplocephalus tutus and the Diversity of Ankylosaurid Dinosaurs in the Late Cretaceous of Alberta, Canada, and Montana, USA", PLoS ONE 8(5): e62421. doi:10.1371/journal.pone.0062421
Noten
- ↑ Euoplocephalus. www.prehistoric-wildlife.com. Gearchiveerd op 22 december 2022. Geraadpleegd op 22-12-2022.
- ↑ Fossilworks: Euoplocephalus. www.fossilworks.org. Geraadpleegd op 22-12-2022.