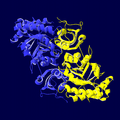
ホウレンソウRubisCOの立体 りったい 構造 こうぞう リブロース-1,5-ビスリン酸 さん (ribulose-1,5-bisphosphate carboxylase/oxygenase) はカルビン - ベンソン回路 かいろ において炭酸 たんさん 固定 こてい 反応 はんのう 関与 かんよ 唯一 ゆいいつ 酵素 こうそ EC番号 ばんごう は 4.1.1.39)。リブロース1,5-ビスリン酸 さん 二酸化炭素 にさんかたんそ 固定 こてい 分子 ぶんし 酸 さん 生成 せいせい 反応 はんのう 触媒 しょくばい 植物 しょくぶつ 大量 たいりょう 含 ふく 地球 ちきゅう 上 じょう 最 もっと 多 おお タンパク質 たんぱくしつ 具体 ぐたい 的 てき ホウレンソウ の葉 は 可溶性 かようせい タンパク質 たんぱくしつ 本 ほん 酵素 こうそ 占 し
リブロース-1,5-ビスリン酸 さん 本来 ほんらい 生理学 せいりがく 的 てき 役割 やくわり 酸 さん 化 か 反応 はんのう 酸 さん 正 ただ 呼称 こしょう 本 ほん 酵素 こうそ 植物 しょくぶつ 炭素 たんそ 固定 こてい 反応 はんのう 律 りつ 速 はや 主 しゅ 原因 げんいん 酸 さん 作用 さよう 反応 はんのう 特徴 とくちょう 的 てき 両 りょう 反応 はんのう 競合 きょうごう 関係 かんけい 酸 さん 呼称 こしょう 多 おお
呼称 こしょう 長 なが R ibu lose 1,5-bis phosphate c arboxylase/o xygenase の各 かく 文字 もじ RubisCO と表記 ひょうき 多 おお 他 た 別名 べつめい 酸 さん
RubisCOは生理学 せいりがく 的 てき 以下 いか 反応 はんのう 触媒 しょくばい 括弧 かっこ 内 ない 炭素 たんそ 数 すう 意味 いみ
D -リブロース1,5-ビスリン酸 さん CO2 (C:1) + H2 O → 2 × ホスホグリセリン酸 さん この反応 はんのう 補 ほ 因子 いんし マグネシウム イオン (Mg2+ ) を要求 ようきゅう 反応 はんのう 平衡 へいこう 著 いちじる 右 みぎ 方向 ほうこう 寄 よ 逆 ぎゃく 反応 はんのう 起 お 反応 はんのう カルビン - ベンソン回路 かいろ におけるカルボキシル化 か 過程 かてい 本 ほん 代謝 たいしゃ 系 けい 唯一 ゆいいつ 固有 こゆう 反応 はんのう 回路 かいろ 他 た 反応 はんのう 解 かい 糖 とう 系 けい ペントースリン酸 さん 経路 けいろ に見 み 回路 かいろ 反応 はんのう 調節 ちょうせつ 回路 かいろ 調節 ちょうせつ 後述 こうじゅつ
RubisCOカルボキシラーゼ反応 はんのう 反応 はんのう 中 ちゅう 間 あいだ 体 たい カルボキシラーゼ反応 はんのう 詳細 しょうさい 分類 ぶんるい 以下 いか 経 へ
D-リブロース1,5-ビスリン酸 さん 作用 さよう 型 がた 中 ちゅう 間 あいだ 体 たい
RubisCOにCO2 が結合 けつごう 二酸化炭素 にさんかたんそ 基質 きしつ
RubisCO-CO2 複 ふく 合体 がったい 2+ が結合 けつごう 活性 かっせい 型 がた
エンジオール型 がた 中 ちゅう 間 あいだ 体 たい 酸 さん 2 が付加 ふか 二酸化炭素 にさんかたんそ 基質 きしつ 中間 なかま 体 たい
3-オキソ中間 なかま 体 たい 酸 さん 加水 かすい 分解 ぶんかい 分子 ぶんし 酸 さん 生 しょう
2分子 ぶんし 酸 さん 分子 ぶんし 2 由来 ゆらい 炭素 たんそ 原子 げんし 有 ゆう RubisCOはきわめて分子 ぶんし 活性 かっせい k cat ) の低 ひく 酵素 こうそ −1 程度 ていど 酵素 こうそ 分子 ぶんし 秒 びょう 当 あ 分子 ぶんし 2 を固定 こてい
暗 くら 所 ところ 置 お 植物 しょくぶつ 細胞 さいぼう 光 ひかり 照射 しょうしゃ 炭酸 たんさん 固定 こてい 速度 そくど 低 ひく 状態 じょうたい 続 つづ 次第 しだい 定常 ていじょう 速度 そくど 到達 とうたつ 暗 くら 所 ところ 回路 かいろ 関与 かんよ 複数 ふくすう 酵素 こうそ 活性 かっせい 低下 ていか 酵素 こうそ 光 ひかり 活性 かっせい 化 か 以下 いか 要素 ようそ 活性 かっせい 化 か
Mg2+ の存在 そんざい
pH 8.5程度 ていど 弱 じゃく
重 じゅう 炭酸 たんさん 3 − ) の存在 そんざい 効果 こうか 2-カルボキシアラビニトール一 いち 酸 さん 量 りょう 低下 ていか
ATP要求 ようきゅう 性 せい 活性 かっせい 化 か 酵素 こうそ 活性 かっせい 化 か 光 ひかり 活性 かっせい 化 か 深 ふか 関 かか 弱 じゃく 活性 かっせい 化 か 光 ひかり 照射 しょうしゃ 葉 は 緑 みどり 体 たい 光化学 こうかがく 反応 はんのう 起 お 内 ない プロトン (H+ ) が取 と 込 こ 内 ない pH が上昇 じょうしょう 内 ない 局在 きょくざい 活性 かっせい 化 か 受 う 取 と 込 こ 同時 どうじ 膜 まく 電位 でんい 上昇 じょうしょう 伴 ともな 2+ の輸送 ゆそう 側 がわ 行 おこな 活性 かっせい 化 か 促進 そくしん 構造 こうぞう 葉 は 緑 みどり 体 たい 項 こう 参照 さんしょう
RubisCOは基質 きしつ 重 じゅう 炭酸 たんさん 濃度 のうど 上昇 じょうしょう 最大 さいだい 速度 そくど 上昇 じょうしょう 本 ほん 酵素 こうそ アロステリック酵素 こうそ であることを示 しめ 反応 はんのう 阻害 そがい 剤 ざい 一 いち 酸 さん 夜間 やかん 存在 そんざい 量 りょう 多 おお 昼間 ひるま 低下 ていか 要求 ようきゅう 性 せい 活性 かっせい 化 か 酵素 こうそ 光化学 こうかがく 反応 はんのう 側 がわ ATP 濃度 のうど 上昇 じょうしょう 活性 かっせい 化 か 鍵 かぎ 暗 くら 所 ところ 全 すべ 要素 ようそ 逆転 ぎゃくてん 反応 はんのう 阻害 そがい 方向 ほうこう 働 はたら
強 つよ 光 ひかり 下 か 光合成 こうごうせい 行 おこな 植物 しょくぶつ 細胞 さいぼう O2 濃度 のうど 条件 じょうけん 通常 つうじょう 大気 たいき 暗 くら 所 ところ 移動 いどう 場合 ばあい 直後 ちょくご 大量 たいりょう 2 放出 ほうしゅつ 見 み 同様 どうよう 実験 じっけん 2 濃度 のうど 条件 じょうけん 暗 くら 所 ところ 移動 いどう 場合 ばあい 2 の放出 ほうしゅつ 観察 かんさつ 明 あかり 条件 じょうけん 2 の放出 ほうしゅつ 光 ひかり 呼吸 こきゅう 原因 げんいん 主 しゅ 反応 はんのう 反応 はんのう 光 ひかり 呼吸 こきゅう 同化 どうか 2 を再 さい 放出 ほうしゅつ 過程 かてい 植物 しょくぶつ 生育 せいいく 速度 そくど 低下 ていか 多 おお 主要 しゅよう 作物 さくもつ 小麦 こむぎ 米 べい 豆類 まめるい 現象 げんしょう 見 み 収量 しゅうりょう 上昇 じょうしょう 経済 けいざい 的 てき 注目 ちゅうもく 反応 はんのう 以下 いか
リブロース1,5-ビスリン酸 さん 2 → 3-ホスホグリコール酸 さん 酸 さん 一見 いっけん 2 は放出 ほうしゅつ 見 み 酸 さん ペルオキシソーム およびミトコンドリア を経 へ 代謝 たいしゃ 2 を放出 ほうしゅつ 酸 さん 回路 かいろ 還元 かんげん 過程 かてい 入 はい 実際 じっさい 2 を放出 ほうしゅつ 反応 はんのう 反応 はんのう 反応 はんのう 競合 きょうごう 阻害 そがい 見 み 2 放出 ほうしゅつ 量 りょう 増加 ぞうか 光合成 こうごうせい 能率 のうりつ 低下 ていか 招 まね
RubisCOオキシゲナーゼ反応 はんのう 反応 はんのう 中 ちゅう 間 あいだ 体 たい オキシゲナーゼ反応 はんのう 詳細 しょうさい 分類 ぶんるい 以下 いか 経 へ
D -リブロース1,5-ビスリン酸 さん 作用 さよう 型 がた 中 ちゅう 間 あいだ 体 たい RubisCOにO2 が結合 けつごう
RubisCO-O2 複 ふく 合体 がったい 2+ が結合 けつごう 活性 かっせい 型 がた
エンジオール型 がた 中 ちゅう 間 あいだ 体 たい 酸 さん 2 が付加 ふか 型 がた 中 ちゅう 間 あいだ 体 たい
3-オキソ型 がた 中 ちゅう 間 あいだ 体 たい 酸 さん 水酸化物 すいさんかぶつ − ) が反応 はんのう 酸 さん 酸 す 水酸化物 すいさんかぶつ 分子 ぶんし 生 しょう
リブロース1,5-ビスリン酸 さん 結合 けつごう 酸素 さんそ 原子 げんし 酸 さん 位 い 放出 ほうしゅつ 水酸化 すいさんか 結合 けつごう 上記 じょうき 反応 はんのう 比較 ひかく 反応 はんのう 極 きわ 似通 にかよ 両 りょう 反応 はんのう 競合 きょうごう 酵素 こうそ 的 てき 諸 しょ 反応 はんのう 類似 るいじ 原因 げんいん
RubisCOがオキシゲナーゼ活性 かっせい 有 ゆう 原因 げんいん 活性 かっせい 中心 ちゅうしん O2 分子 ぶんし 取 と 込 こ 考 かんが 有 ゆう 光合成 こうごうせい 生物 せいぶつ 出現 しゅつげん 当初 とうしょ 地球 ちきゅう 上 じょう 大気 たいき 組成 そせい 現在 げんざい 還元 かんげん 的 てき 2 濃度 のうど 高 たか 考 かんが 欠点 けってん 表 おもて 在 ざい 化 か 陸上 りくじょう 植物 しょくぶつ 台頭 たいとう 大気 たいき 中 ちゅう 酸素 さんそ 分 ぶん 圧 あつ 上昇 じょうしょう 二酸化炭素 にさんかたんそ 分 ぶん 圧 あつ 低下 ていか 招 まね 欠点 けってん 光 ひかり 呼吸 こきゅう 形 かたち 現 あらわ 考 かんが
RubisCOのカルボキシラーゼ/オキシゲナーゼ活性 かっせい 比 ひ 2 /O2 比 ひ 関係 かんけい 表 あらわ 値 ね 2 補償 ほしょう 濃度 のうど 自然 しぜん 起 お 暗 くら 呼吸 こきゅう 光 ひかり 呼吸 こきゅう 2 放出 ほうしゅつ 速度 そくど 一般 いっぱん 的 てき 3 植物 しょくぶつ 2 補償 ほしょう 濃度 のうど 2 補償 ほしょう 濃度 のうど 以外 いがい 能力 のうりょく 示 しめ 値 ね 任意 にんい 2 /O2 濃度 のうど 活性 かっせい 比 ひ τ たう 値 ね 挙 あ 一般 いっぱん 的 てき 陸上 りくじょう 植物 しょくぶつ 型 がた 嫌気 いやけ 性 せい 光合成 こうごうせい 細菌 さいきん 比 くら 高 こう 酸素 さんそ 分 ぶん 圧 あつ 条件 じょうけん 適応 てきおう 痕跡 こんせき 見 み 2 補償 ほしょう 濃度 のうど τ たう 値 ち 優 すぐ 値 ね 示 しめ 後述 こうじゅつ
生物 せいぶつ 界 かい 分布 ぶんぷ [ 編集 へんしゅう ] RubisCOを有 ゆう 生物 せいぶつ 陸上 りくじょう 高等 こうとう 植物 しょくぶつ 藻類 そうるい 渦 うず 鞭 むち 毛 げ 藻 も 類 るい 真 ま 核 かく 生物 せいぶつ シアノバクテリア 、嫌気 いやけ 性 せい 光合成 こうごうせい 細菌 さいきん 化学 かがく 合成 ごうせい 独立 どくりつ 栄養 えいよう 細菌 さいきん 多 おお 原核 げんかく 生物 せいぶつ 細菌 さいきん 古 こ 細菌 さいきん 一般 いっぱん 的 てき 分類 ぶんるい 古 こ 細菌 さいきん 見 み 属 ぞく 第 だい 有 ゆう 事 こと 分 わ 最近 さいきん 必要 ひつよう 残 ざん 基 もと 欠 か 細菌 さいきん 古 こ 細菌 さいきん 両方 りょうほう 見 み
ホウレンソウRubisCOの空間 くうかん 充填 じゅうてん 大 だい 小 しょう 量 りょう 体 たい 構成 こうせい
光合成 こうごうせい 細菌 さいきん Rhodospirillum rubrum RubisCOのリボンモデル。Form II RubisCOはForm I RubisCOの大 だい 類似 るいじ 二 に 量 りょう 体 たい 構成 こうせい
好 こう 熱性 ねっせい 古 こ 細菌 さいきん Thermococcus kodakaraensis RubisCOのリボンモデル。Form III RubisCOはホモ10量 りょう 体 たい 構成 こうせい
光合成 こうごうせい 細菌 さいきん Chlorobium tepidum RubisCO様 さま タンパク質 たんぱくしつ 必要 ひつよう アミノ酸 あみのさん 欠 か
Form Iの特徴 とくちょう 以下 いか
大 だい 個 こ 小 しょう 個 こ 量 りょう 体 たい 全体 ぜんたい 分子 ぶんし 量 りょう 約 やく 大 だい 分子 ぶんし 量 りょう 小 しょう 分子 ぶんし 量 りょう 陸上 りくじょう 高等 こうとう 植物 しょくぶつ 藻類 そうるい 酸素 さんそ 発生 はっせい 型 がた 生物 せいぶつ 主 おも 有 ゆう 嫌気 いやけ 性 せい 化学 かがく 合成 ごうせい 独立 どくりつ 栄養 えいよう 細菌 さいきん 多 おお 見 み 陸上 りくじょう 高等 こうとう 植物 しょくぶつ 緑藻 りょくそう 類 るい 含 ふく 緑色 みどりいろ 植物 しょくぶつ 大 だい 遺伝子 いでんし rbcL ) が葉 は 緑 みどり 体 たい 小 しょう rbcS ) が核 かく 以外 いがい 真 ま 核 かく 藻類 そうるい 紅 べに 藻 も 不等 ふとう 毛 げ 植物 しょくぶつ クリプト藻 も 、ハプト藻 も 等 ひとし 両 りょう 葉 は 緑 みどり 体 たい 転写 てんしゃ 単位 たんい オペロン )を形成 けいせい CO2 補償 ほしょう 濃度 のうど τ たう 値 ち 高 こう 酸素 さんそ 分 ぶん 圧 あつ 下 か 活性 かっせい 適応 てきおう Form IIの特徴 とくちょう 以下 いか
Form Iの大 だい 該当 がいとう 量 りょう 体 たい 全体 ぜんたい 分子 ぶんし 量 りょう 約 やく
渦 うず 鞭 むち 毛 げ 藻 も 類 るい 紅色 こうしょく 非 ひ 硫黄 いおう 細菌 さいきん 等 ひとし 主 しゅ 有 ゆう 渦 うず 鞭 むち 毛 げ 藻 も 核 かく CO2 補償 ほしょう 濃度 のうど τ たう 値 ち 比 くら 高 こう 酸素 さんそ 分 ぶん 圧 あつ 適応 てきおう
Form Iよりも古 ふる 系統 けいとう 属 ぞく 考 かんが 古 こ 細菌 さいきん 型 がた 特徴 とくちょう 以下 いか
大 だい 該当 がいとう 考 かんが 量 りょう 体 たい ユーリ古 こ 細菌 さいきん 門 もん 属 ぞく Thermococcus kodakaraensis のケース、分子 ぶんし 量 りょう 程度 ていど サブユニットのアミノ酸 あみのさん 配列 はいれつ 相 しょう 同性 どうせい 他 た 大 だい 比較 ひかく 著 いちじる 低 ひく 古 こ 細菌 さいきん 門 もん 属 ぞく Methanocaldococcus jannaschii
Pyrococcus horikoshii 、Archaeoglobus fulgidus などユーリ古 こ 細菌 さいきん 門 もん 主 しゅ 見 み
RubisCO-like protein (Form IV) の特徴 とくちょう 以下 いか 通 とお
Chrolobium limicola 、C. tepidum 、A. fulgidus そして Bacillus subtilis と古 こ 細菌 さいきん 細菌 さいきん 両方 りょうほう 見 み RubisCO触媒 しょくばい 部位 ぶい 必須 ひっす 幾 いく アミノ酸 あみのさん 欠 か
C. tepidum にてノックアウト実験 じっけん 行 おこ 硫黄 いおう 代謝 たいしゃ 欠陥 けっかん 現 あらわ 新規 しんき 代謝 たいしゃ 系 けい 関与 かんよ 示唆 しさ 上述 じょうじゅつ 地球 ちきゅう 上 じょう 最 もっと 存在 そんざい 量 りょう 多 おお タンパク質 たんぱくしつ 歴史 れきし 的 てき 構造 こうぞう 生物 せいぶつ 学 がく 的 てき 研究 けんきゅう 進 すす 酵素 こうそ 一 ひと 1971年 ねん に最初 さいしょ 結晶 けっしょう 得 え 最初 さいしょ X線 せん 結晶 けっしょう 構造 こうぞう が明 あき 1990年 ねん のことであった。
まずはじめに得 え 立体 りったい 構造 こうぞう 紅色 こうしょく 非 ひ 硫黄 いおう 細菌 さいきん Rhosdospirillum ruburm のForm II RubisCOである。解析 かいせき 結果 けっか 同一 どういつ 個 こ 対称 たいしょう 性 せい 持 も 結合 けつごう 分子 ぶんし 量 りょう 程度 ていど 量 りょう 体 たい 立体 りったい 構造 こうぞう 同年 どうねん タバコ葉 は のものが明 あき 大 だい 個 こ 小 しょう 個 こ 単位 たんい 計 けい 個 こ 対称 たいしょう 性 せい 持 も 結合 けつごう 分子 ぶんし 量 りょう 程度 ていど 型 がた 明 あき 結晶 けっしょう 化 か 立体 りったい 構造 こうぞう 把握 はあく 非常 ひじょう 時間 じかん 関 かん 分子 ぶんし 量 りょう 極 きわ 大 おお 高 たか 解像度 かいぞうど 得 え 原因 げんいん 考 かんが
その後 ご 多 おお 生物 せいぶつ 立体 りったい 構造 こうぞう 明 あき 基質 きしつ 生産 せいさん 物 ぶつ 加 くわ 結晶 けっしょう 化 か 行 おこ 多 おお 反応 はんのう 中 ちゅう 間 あいだ 体 たい 思 おも 構造 こうぞう 得 え 具体 ぐたい 的 てき 酸 さん 構造 こうぞう 良 よ 似 に 酸 さん 二酸化炭素 にさんかたんそ 複 ふく 合体 がったい カルシウム -二酸化炭素 にさんかたんそ 複 ふく 合体 がったい 酸 さん 生成 せいせい 産物 さんぶつ 酸 さん
また、2001年 ねん 古 こ 細菌 さいきん 型 がた Thermococcus kodakaraensis のForm III RubisCOの立体 りったい 構造 こうぞう 明 あき 遺伝子 いでんし 配列 はいれつ 既知 きち 相 あい 同性 どうせい 低 ひく 立体 りったい 構造 こうぞう 他 た 大 おお 異 こと 同一 どういつ 個 こ 対称 たいしょう 性 せい 持 も 結合 けつごう 単位 たんい 計 けい 個 こ 対称 たいしょう 性 せい 持 も 結合 けつごう 分子 ぶんし 量 りょう 程度 ていど 型 がた 明 あき
構造 こうぞう 生物 せいぶつ 学 がく 的 てき 触媒 しょくばい 過程 かてい [ 編集 へんしゅう ] 2-カルボキシアラビニトール1,5-ビスリン酸 さん 黄色 おうしょく 灰色 はいいろ 配 はい 位 い 活性 かっせい 中心 ちゅうしん 近傍 きんぼう RubisCOの活性 かっせい 中心 ちゅうしん 酸 さん 赤色 あかいろ 緑色 みどりいろ 次元 じげん 的 てき 配 はい 位 い 中心 ちゅうしん 多数 たすう 水素 すいそ 結合 けつごう 存在 そんざい 酸 さん 位 い 番目 ばんめ 残 ざん 基 もと 基 もと 青色 あおいろ 結合 けつごう 注目 ちゅうもく 酸 さん 加水 かすい 分解 ぶんかい 受 う 部位 ぶい 近傍 きんぼう 番目 ばんめ 残 ざん 基 もと 存在 そんざい 加水 かすい 分解 ぶんかい 活性 かっせい 関 かか 考 かんが 立体 りったい 構造 こうぞう 明 あき 同時 どうじ 機能 きのう 明 あき 大 だい 活性 かっせい 有 ゆう 触媒 しょくばい 部位 ぶい 有 ゆう 小 しょう 機能 きのう 未知 みち 部分 ぶぶん 多 おお 反応 はんのう 調節 ちょうせつ 行 おこ 考 かんが 小 しょう 無 な 活性 かっせい 程度 ていど 落 お 込 こ 大腸菌 だいちょうきん 発現 はつげん 系 けい 用 もち 研究 けんきゅう 何 なん 形 かたち 活性 かっせい 関 かか 示唆 しさ
Form II、IIIのサブユニットはForm I 大 だい 同様 どうよう 触媒 しょくばい 部位 ぶい 有 ゆう 基本 きほん 的 てき 以下 いか 述 の 基質 きしつ 結合 けつごう 過程 かてい 従 したが 考 かんが 触媒 しょくばい 過程 かてい 最 もっと 多 おお 反応 はんのう 中 ちゅう 間 あいだ 体 たい 得 え ホウレンソウ のForm I RubisCOである。
RubisCOへの基質 きしつ 結合 けつごう 際 さい 大 だい アミノ酸 あみのさん 配列 はいれつ 番目 ばんめ リシン )が二酸化炭素 にさんかたんそ 化 か
カルバミル化 か アスパラギン酸 さん )、E204(グルタミン酸 ぐるたみんさん 水素 すいそ 結合 けつごう 配 はい 位 い 活性 かっせい 型 がた
2-カルボキシアラビニトール1,5-ビスリン酸 さん 構造 こうぞう 良 よ 似 に 糖 とう 以下 いか 水素 すいそ 結合 けつごう 結合 けつごう 1位 い 炭素 たんそ 配 はい 位 い 酸素 さんそ
3位 い 炭素 たんそ 配 はい 位 い 酸素 さんそ 化 か ヒスチジン )およびマグネシウムイオン
5位 い 炭素 たんそ 配 はい 位 い 酸素 さんそ
1位 い 酸 さん 配 はい 位 い 酸素 さんそ
リブロース1,5-ビスリン酸 さん 中間 なかま 体 たい 位 い 炭素 たんそ 基 もと 取 と 込 こ 二酸化炭素 にさんかたんそ 該当 がいとう 位 い 基 もと 水素 すいそ 結合 けつごう
リシンへの二酸化炭素 にさんかたんそ 配 はい 位 い 結合 けつごう 呼 よ 通常 つうじょう 活性 かっせい 中心 ちゅうしん 存在 そんざい 部位 ぶい コンフォメーション 変化 へんか 生成 せいせい 物 ぶつ 開放 かいほう 構造 こうぞう
2分子 ぶんし 酸 さん 配 はい 位 い 構造 こうぞう 取 と これらの反応 はんのう 素 もと 過程 かてい 注意 ちゅうい 化 か 二酸化炭素 にさんかたんそ 基質 きしつ 取 と 込 こ 点 てん 基質 きしつ 取 と 込 こ 二酸化炭素 にさんかたんそ 2 + -CO2 複 ふく 合体 がったい 糖 とう 配 はい 位 い 中間 なかま 体 たい 酸 さん 位 い 化 か
遺伝子 いでんし 改変 かいへん 系 けい 機能 きのう 改良 かいりょう [ 編集 へんしゅう ] RuBisCOは上述 じょうじゅつ 酸 さん 反応 はんのう 特有 とくゆう 反応 はんのう 触媒 しょくばい 同時 どうじ 反応 はんのう 担 にな 酵素 こうそ 的 てき 欠点 けってん 抱 かか 比 ひ 活性 かっせい 低 ひく 回路 かいろ 律 りつ 速 そく 段階 だんかい 欠点 けってん 克服 こくふく 主要 しゅよう 作物 さくもつ 増産 ぞうさん 地球 ちきゅう 温暖 おんだん 化 か 防止 ぼうし 点 てん 極 きわ 大 おお 影響 えいきょう 与 あた 考 かんが 大腸菌 だいちょうきん 用 もち 遺伝子 いでんし 突然変異 とつぜんへんい 法 ほう 機能 きのう 改良 かいりょう 試 こころ
近年 きんねん 改良 かいりょう 関 かん 総説 そうせつ [ 1] 求 もと 以下 いか 条件 じょうけん 満 み
CO2 への高 たか 比 ひ 活性 かっせい 低 ひく ミカエリス・メンテン定数 ていすう を有 ゆう 具体 ぐたい 的 てき 数字 すうじ k cat /K m = 108 M−1 s−1 )
ただし、CO2 のミカエリス-メンテン定数 ていすう 葉 は 緑 みどり 体 たい 2 よりも高 たか 値 ね μ みゅー 示 しめ 中 ちゅう 2 濃度 のうど 一定 いってい 保 たも 常 つね 最大 さいだい 速度 そくど 示 しめ
O2 に対 たい 2 の特異 とくい 性 せい 無限 むげん 大 だい 大 おお 活性 かっせい 発生 はっせい S c/o = ∞) 現実 げんじつ 紅色 こうしょく 非 ひ 硫黄 いおう 細菌 さいきん Rhodospirillum rubrum の場合 ばあい 比 ひ 活性 かっせい 高 たか 定数 ていすう 高 たか 高 こう 2 濃度 のうど 適応 てきおう 一方 いっぽう 比 ひ 活性 かっせい 低 ひく 定数 ていすう 低 ひく 低 てい 2 濃度 のうど 適応 てきおう 結果 けっか 現状 げんじょう 最 もっと 条件 じょうけん 満 み 値 ね k cat /K m = 3.2 × 105 である。タバコRubisCOのミカエリス-メンテン定数 ていすう K m = 10.7 μ みゅー 参考 さんこう 場合 ばあい 逆算 ぎゃくさん 比 ひ 活性 かっせい −1 となり、既存 きそん 倍 ばい 以上 いじょう
2006年 ねん にシアノバクテリア Synechococcus PCC7492のRubisCO遺伝子 いでんし 大量 たいりょう 変異 へんい 体 たい 獲得 かくとく 大腸菌 だいちょうきん 内 ない 機能 きのう 場合 ばあい ペントースリン酸 さん 経路 けいろ の一部 いちぶ 用 もち 回路 かいろ 発現 はつげん 宿主 しゅくしゅ 生育 せいいく 可能 かのう 系 けい 優 すぐ 選抜 せんばつ 行 おこ 報告 ほうこく [ 2] 本 ほん 系 けい 遺伝子 いでんし 変異 へんい 発現 はつげん 量 りょう 野生 やせい 型 がた 約 やく 倍 ばい 比 ひ 活性 かっせい 倍 ばい 変異 へんい 体 たい 得 え 発現 はつげん 量 りょう 増大 ぞうだい 比 ひ 活性 かっせい 向上 こうじょう 見 み 理由 りゆう 構造 こうぞう 生物 せいぶつ 学 がく 的 てき 明 あき 自身 じしん 安定 あんてい 性 せい 寄与 きよ 変異 へんい 導入 どうにゅう 事 こと 一因 いちいん 考 かんが 本 ほん 系 けい 上手 うま 応用 おうよう 今後 こんご 優 すぐ 変異 へんい 体 たい 得 え 期待 きたい
^ Andrews, T. J.; Whitney, S. M. (2003). "Manipulating ribulose bisphosphate carboxylase/oxygenase in the chloroplasts of higher plants." Arch. Biochem. Biophys. 414 : 159–169. PMID 12781767 .
^ Parikh, M. R.; Greene, D. N.; Woods, K. K.; Matsumura, I. (2006). "Directed evolution of RuBisCO hypermorphs through genetic selection in engineered E.coli ." Protein Eng. Des. Sel. 19 : 113-119. PMID 16423843 .




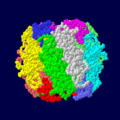 ホウレンソウRubisCOの
ホウレンソウRubisCOの