恐竜
Dinosauria | |||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||||||||||||||||||||||||||||||
( | |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| Dinosauria Owen, 1842 | |||||||||||||||||||||||||||||||||||||||||||||
| Dinosaur | |||||||||||||||||||||||||||||||||||||||||||||
概要
名称

オーウェンはδεινός の
定義
古典 的 定義
分岐 学 的 定義
この
| |||||||||||||||||||||||||||||||||||||||||||

よって
また、より
これらの
なお、
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||

特徴
特徴 的 な派生 形質

(
後前 頭骨 (postfrontal)を二 次 的 に失 う。上腕 骨 の三角 筋 稜 (delto-pectoral crest)が発達 する。腸 骨 、恥骨 、坐骨 で構成 された骨盤 に、大腿 骨 がはまり込 む場所 である寛 骨 臼 (かんこつきゅう)が貫通 している。仙 椎 を構成 する骨 が3個 以上 (ワニやトカゲなど、多 くの爬虫類 ではより少 ないことが多 い)。脛骨 前面 に距骨突起 が成立 する。
など。
これらのいくつかは、
進化 史
登場 と初期 進化
非 鳥類 型 恐竜 の絶滅
K-Pg
しかし
鳥類
生物 学 的 特徴

その
姿勢 ・歩行
この
カメは
肌 ・色

2008
羽毛

そして1990
気嚢 式 呼吸

ペルム
恒温 性
しかしながら
またエドウィン・ハリス・コルバートらがアメリカアリゲーターの
生態 ・社会 性

また、
繁殖 ~孵化
獣 脚 類
竜 脚 類 (主 にティタノサウルス形 類 )
鳥 脚 類
研究 史
発見


1815
1822
1842
化石 戦争
鳥 との関係

1861
恐竜 ルネッサンス
1969
2000年代 以降 の主 な報告
指 の謎
これに
軟組織 の発見
2000
別種 か、それとも?
トリケラトプスとトロサウルス[77][78]、パキケファロサウルスとスティギモロクとドラコレックスのように、これまで
復元
かつて
下位 分類
なお、




上位 分類
竜 盤 類 Saurischia
[
- ?エオラプトル
科 Eoraptoridae - エオラプトル - ?ヘレラサウルス
類 Herrerasauria- ヘレラサウルス
科 Herrerasauridae - ヘレラサウルス、レヴェルトラプトル - スタウリコサウルス
科 Staurikosauridae - スタウリコサウルス (三 畳 紀 、南米 )
- ヘレラサウルス
- グアイバサウルス
科 Guaibasauridae - グアイバサウルス 新 獣 脚 類 Neotheropoda- ケラトサウルス
下目 Ceratosauria- コエロフィシス
上 科 Coelophysoidea- ゴジラサウルス Gojirasaurus
- コエロフィシス
科 Coelophysidae- プロコンプソグナトゥス
亜 科 Procompsognathinae - プロコンプソグナトゥス - コエロフィシス
亜 科 Coelophysinae - コエロフィシス、サルトプス、メガフォサウルス
- プロコンプソグナトゥス
- ?ハルティコサウルス
科 - ハルティコサウルス、ディロフォサウルス
- ネオケラトサウルス
類 (新 ケラトサウルス類 ) Neoceratosauria
- コエロフィシス
- テタヌラ
下目 Tetanurae- ?クリオロフォサウルス Cryolophosaurus
- メガロサウルス
上 科 Megalosauroidea - アヴェテロポーダ
類 Avetheropoda(新 テタヌラ類 Neotetanurae)- カルノサウルス
類 Carnosauria- アロサウルス
上 科 Allosauroidea- モノロフォサウルス Monolophosaurus
- シンラプトル
科 Sinraptoridae - シンラプトル、ヤンチュアノサウルス、スゼチュアノサウルス - アロサウルス
科 Allosauridae - アロサウルス、クリオロフォサウルス、サウロファガナクス、アクロカントサウルス - カルカロドントサウルス
科 Carcharodontosauridae - カルカロドントサウルス、ギガノトサウルス、マプサウルス
Allosaurus fragilis(アロサウルス・フラギリス)の 爪 のレプリカ
- アロサウルス
- コエルロサウルス
類 Coelurosauria- ?オルニトレステス
科 Ornitholestidae -オルニトレステス - コンプソグナトゥス
科 Compsognathidae - コンプソグナトゥス、シノサウロプテリクス(中華 竜 鳥 ) - コエルルス
科 Coeluridae - コエルルス、オルニトレステス - ティラノサウルス
上 科 Tyrannosauroidea-エオティラヌス、 - オルニトミモサウルス
類 Ornithomimosauria- オルニトミムス
科 Ornithomimidae - オルニトミムス、ストルティオミムス、ガリミムス
マニラプトル 類 の一種 、ヴェロキラプトルの復元 。
- オルニトミムス
- マニラプトル
類 Maniraptora- アルヴァレスサウルス
科 Alvarezsauridae - モノニクス、アルヴァレスサウルス、パタゴニクス - オヴィラプトロサウルス
形 類 Oviraptoriformes- オヴィラプトロサウルス
類 Oviraptorosauria- アビミムス
科 Avimimidae - アビミムス - カウディプテリクス
科 Caudipteridae - カウディプテリクス - カエナグナトゥス
上 科 Caenagnathoidea- カエナグナトゥス
科 Caenagnathidae - カエナグナタシア、キロステノテス、エルミサウルス - オヴィラプトル
科 Oviraptoridae - オヴィラプトル、アジャンキンゲニア、インキシヴォサウルス
- カエナグナトゥス
- アビミムス
- テリジノサウルス
類 Therizinosauria- テリジノサウルス
上 科 Therizinosauroidea
- テリジノサウルス
- オヴィラプトロサウルス
原 鳥類 Paraves- ペドペンナ Pedopenna
- ?スカンソリオプテリクス
科 Scansoriopterygidae - スカンソリオプテリクス、エピデクシプテリクス - デイノニコサウルス
類 (デイノニクス竜 類 )Deinonychosauria 鳥類 Aves(現生 鳥類 の先祖 )
- アルヴァレスサウルス
- ?オルニトレステス
- カルノサウルス
- ケラトサウルス


- テコドントサウルス
科 Thecodontosauridae - テコドントサウルス 原 竜 脚下 目 Prosauropoda- ?アンキサウルス
科 Anchisauridae - アンキサウルス - プラテオサウルス
科 Plateosauridae - プラテオサウルス、ウナイサウルス - メラノロサウルス
科 Melanorosauridae - メラノロサウルス - マッソスポンディルス
科 Massospondylidae - マッソスポンディルス、ルーフェンゴサウルス
- ?アンキサウルス
竜 脚下 目 Sauropoda- ?ヴルカノドン
科 Vulcanodontidae - ヴルカノドン 真 竜 脚 亜 目 (エウサウロポーダ類 ) Eusauropoda- ?イービノサウルス
- マメンチサウルス
類 - マメンチサウルス
科 Mamenchisauridae - マメンチサウルス、チュアンジェサウルス - ケティオサウルス
類 - ケティオサウルス
科 Cetiosauridae - ケティオサウルス、ロエトサウルス - ジョバリア
科 - ジョバリア - オメイサウルス
科 Omeisauridae - オメイサウルス 新 竜 脚 類 Neosauropoda- ハプロカントサウルス Haplocanthosaurus
- ディプロドクス
類 Diplodocimorpha- レッバキサウルス
科 Rebbachisauridae - ディプロドクス
上 科 Diplodocoidea- Flagellicaudata
- ディプロドクス
科 Diplodocidae - ディクラエオサウルス
科 Dicraeosauridae - ディクラエオサウルス、アマルガサウルス、レッバキサウルス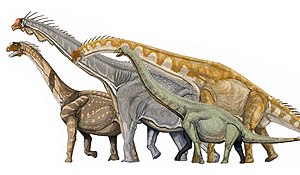
代表 的 なマクロナリア類 の復元 。左 からカマラサウルス、ブラキオサウルス、ギラファティタン、エウヘロプス
- ディプロドクス
- Flagellicaudata
- レッバキサウルス
- マクロナリア
類 Macronaria- カマラサウルス
類 Camarasauromorpha- カマラサウルス
科 Camarasauridae - カマラサウルス - ティタノサウルス
形 類 Titanosauriformes- ?アンデサウルス
科 - アンデサウルス、マラウィサウルス - ブラキオサウルス
科 Brachiosauridae - ブラキオサウルス - ティタノサウルス
上 科 Titanosauria
- ?アンデサウルス
- カマラサウルス
- カマラサウルス
- ケティオサウルス
- マメンチサウルス
- ?ヴルカノドン

鳥 盤 類 Ornithischia
ファブロサウルス
ゲナサウリス
装 楯 亜 目 Thyreophora- スクテロサウルス
科 - スクテロサウルス - ユーリ
脚 類 (寛 脚 類 ) Eurypoda- スケリドサウルス
科 Scelidosauridae - スケリドサウルス - ステゴサウルス
類 (剣 竜 下目 ) Stegosauria 曲 竜 下目 (鎧 竜 下目 ) Ankylosauria- ステゴペルタ
族 Stegopeltini - グリプトドントペルタ、ステゴペルタ - ノドサウルス
科 Nodosauridae - ノドサウルス、サウロペルタ、エドモントニア、ドラコペルタ - ポラカントゥス
科 Polacanthidae - ポラカントゥス、ガーゴイロサウルス、ガストニア - アンキロサウルス
科 Ankylosauridae - アンキロサウルス、エウオプロケファルス、サイカニア
- ステゴペルタ
- スケリドサウルス
- スクテロサウルス
角 脚 類 Ceraopoda(新 鳥 盤 類 Neornithischia)- ヘテロドントサウルス
科 Heterodontosauridae - ヘテロドントサウルス 鳥 脚 亜 目 Ornithopoda- ゼフィロサウルス
科 Zephyrosauridae - ゼフィロサウルス - オスニエリア
科 Othnieliidae - オスニエリア 新 鳥 脚 類 Neornithopoda(正 鳥 脚下 目 Euornithopoda)- ヒプシロフォドン
科 Hypsilophodontidae - ヒプシロフォドン - イグアノドン
類 Iguanodontia- ムッタブラサウルス Muttaburrasaurus
真 イグアノドン類 Euiguanodontia- Rhabdodontidae - ラブドドン
- ドリオサウルス
科 Dryosauridae - ドリオサウルス、ヴァルドサウルス - アンキロポレクシア Ankylopollexia
- カンプトサウルス
科 Camptosauridae - カンプトサウルス、ドラコニクス - イグアノドン
上 科 Iguanodontoidea- イグアノドン
科 Iguanodontidae - イグアノドン、オウラノサウルス、フクイサウルス - プロバクトロサウルス
科 Probactrosauridae - プロバクトロサウルス - ハドロサウルス
科 Hadrosauridae - ハドロサウルス、アナトティタン、コリトサウルス、ランベオサウルス、パラサウロロフス、マイアサウラ、シャントゥンゴサウルス
- イグアノドン
- カンプトサウルス
- ヒプシロフォドン
- ゼフィロサウルス
周 飾 頭 亜 目 Marginocephalia- ステノペリクス Stenopelix
- ヘテロドントサウルス
科 Heterodontosauridae - ヘテロドントサウルス、ゲラノサウルス、ラナサウルス 堅 頭 竜 下目 Pachycephalosauria- ホマロケファレ
上 科 Homalocephaloidea- パキケファロサウルス
科 Pachycephalosauridae - パキケファロサウルス、ステゴケラス - ホマロケファレ
科 Homarocephalidae - ホマロケファレ
- パキケファロサウルス
- ホマロケファレ
角 竜 下目 Ceratopia- プシッタコサウルス
科 Psittacosauridae - プシッタコサウルス 新 角 竜 類 Neoceratopsia
- プシッタコサウルス
- ヘテロドントサウルス
主 な恐竜 の分類 群 の時間 範囲
ホルツによる

日本 で発掘 された主 な化石
北海道 芦別 町 …ティラノサウルス類 北海道 小平 町 …ハドロサウルス類 の後 ろ足 北海道 夕張 市 …ノドサウルス類 の頭骨 の一部 北海道 むかわ町 …ハドロサウルス科 、カムイサウルス・ジャポニクス(むかわ竜 )[86]岩手 県 岩泉 町 …竜 脚 類 (通称 モシリュウ)福島 県 南 相馬 市 …足跡 (ジュラ紀 )福島 県 広野 町 …鳥 脚 類 (通称 ヒロノリュウ)、ティラノサウルス類 (通称 フタバリュウ)[87]福島 県 いわき市 …竜 脚 類 (通称 ヒサノハマリュウ)、鳥 脚 類 群馬 県 神 流 町 (旧 中里 村 )…スピノサウルス類 の歯 、オルニトムス類 (通称 サンチュウリュウ)、獣 脚 類 、足跡 長野 県 小谷 村 …足跡 (ジュラ紀 )富山 県 富山 市 (旧 大山 町 )…イグアノドン類 の歯 、獣 脚 類 、足跡 岐阜 県 白川 町 …竜 脚 類 、足跡 岐阜 県 高山市 (旧 荘川 村 )…イグアノドン類 、ヒプシロフォドン類 の歯 、鳥 脚 類 、獣 脚 類 石川 県 白山 市 (旧 白峰 村 )…竜 脚 類 (通称 ハクサンリュウ)、獣 脚 類 (通称 オピラプトロサウルス類 )、イグアノドン類 (通称 シマリュウ)、アルバロフォサウルス・ヤマグチオロウム(角 脚 類 )[88]、ティラノサウルス類 (通称 カガリュウ)[89]、足跡 福井 県 勝山 市 …フクイサウルス・テトリエンシス(イグアノドン類 )[90]、フクイラプトル・キタダニエンシス(メガラプトル類 )[91]、フクイティタン・ニッポネンシス(竜 脚 類 )[92]、コシサウルス・カツヤマ(イグアノドン類 )[93]、フクイベナートル・パラドクサス(コエルロサウルス類 )、フクイプテリクス・プリマ (鳥 群 )、ティラノミムス・フクイエンシス (オルニトミモサウルス類 )[94]、獣 脚 類 、鳥 脚 類 、足跡 福井 県 大野 市 (旧 和泉 村 )…獣 脚 類 の歯 、足跡 三重 県 鳥羽 市 …ティタノサウルス類 (通称 トバリュウ)、足跡 兵庫 県 丹波 市 .丹波 篠山 市 …タンバティタニス・アミキティアエ(ティタノサウルス形 類 )、ヒプノヴェナトル・マツバラエトオオエオルム(コエルロサウルス類 )、ササヤマグノームス・サエグサイ(新 角 竜 類 )、角 竜 類 、ティラノサウルス類 、鎧 竜 類 など兵庫 県 洲本 市 …ヤマトサウルス・イザナギイ(ハドロサウルス類 )ランベオサウルス類 徳島 県 勝浦 町 …イグアノドン類 の歯 山口 県 下関 市 吉母 …足跡 福岡 県 北九州 市 …獣 脚 類 福岡 県 宮 若 市 (旧 宮田 町 )…獣 脚 類 (ワキノサウルス・サトウイ)長崎 県 長崎 市 …獣 脚 類 [95]熊本 県 御船 町 …ティラノサウルス類 、オルニトミムス類 、テリジノサウルス類 、ドロマエオサウルス類 、アンキロサウルス類 、ハドロサウルス類 、ミフネリュウ、足跡 熊本 県 天草 市 …獣 脚 類 、イグアノドン類 、鳥 脚 類 、竜 脚 類 、足跡 熊本 県 八代 市 …獣 脚 類 (アロサウルス形 類 )、肋骨 [96]鹿児島 県 薩摩 川内 市 (旧 下甑 村 、旧 鹿島 村 )…獣 脚 類 の歯 、イグアノドン類 の歯 、角 竜 類 の歯
博物館
脚注
注釈
出典
- ^ a b
冨田 幸光 ,對比 地 孝 亘 ,三枝 春生 ,池上 直樹 ,平山 廉 ,仲谷 英夫 「恐竜 類 の分岐 分類 におけるクレード名 の和訳 について」『化石 』第 108巻 、日本 古 生物 学会 、2020年 、23-35頁 、doi:10.14825/kaseki.108.0_23。 - ^
巌 佐 庸 ・倉谷 滋 ・斎藤 成 也・塚谷 裕一 編 「生物 分類 表 」『岩波 生物 学 辞典 第 5版 』岩波書店 、2013年 、1531–1666頁 。 - ^
小林 快 次 『恐竜 の学校 』小林 快 次 監修 、ニュートンプレス、2023年 、20頁 。ISBN 978-4-315-52725-4。 - ^ Fassett, J. E.; Zielinski, R. A.; Budahn, J. R. (2002). “Dinosaurs that did not die: evidence for Paleocene dinosaurs in the Ojo Alamo sandstone, San Juan Basin, New Mexico.”. Geological Society of America Special Papers 356: 307-336. doi:10.1130/0-8137-2356-6.307.
- ^ a b Owen, R. (1842). "Report on British Fossil Reptiles." Part II." Report of the Eleventh Meeting of the British Association for the Advancement of Science; Held at Plymouth in July 1841.London: John Murray. pp.: 60-204
- ^ ASIN 0253333490, The Complete Dinosaur ISBN 978-0253333490
- ^ “
恐竜 は「恐 ろしい竜 」ではない”.恐竜 の楽園 (2011年 1月 19日 ). 2012年 12月 10日 時点 のオリジナルよりアーカイブ。2017年 7月 2日 閲覧 。 - ^ “
悩 ましい翻訳 語 ―科学 用語 の由来 と誤訳 ”. はてなブログ. はてな (2011年 6月 14日 ). 2017年 7月 2日 閲覧 。 - ^
小畠 郁生 (監修 )、西川 寛 、高岡 昌江 、三品 隆司 『恐竜 の大 常識 』ポプラ社 、2003年 、14頁 。ISBN 4591078353。 - ^ a b c Dinosauria2nd 2004, pp. [,
要 ページ番号 ], - ^ a b c d e f Fastovsky & Weishampel 2006, pp. [,
要 ページ番号 ], - ^ Benton MJ: Origin and relationships of Dinosauria. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. Zweite Auflage. University of California Press, Berkeley 2004, S. 7-19, ISBN 0-520-24209-2.
- ^ Olshevsky, G. (2000). "An annotated checklist of dinosaur species by continent". Mesozoic Meanderings 3: 1–157.
- ^ Sereno, P. C. (2005). “The logical basis of phylogenetic taxonomy”. Syst. Biol. 54 (4): 595–619. doi:10.1080/106351591007453. PMID 16109704.
- ^ ダレン・ナイシュ、ポール・バレット
著 、小林 快 次 、久保田 克博 、千葉 謙太郎 、田中 康平 監訳 吉田 三知 世 訳 『恐竜 の教科書 最新 研究 で読 み解 く進化 の謎 』創 元 社 、2019年 2月 20日 、45頁 。ISBN 978-4-422-43028-7。 - ^
読売新聞 、2011年 2月 5日 22時 8分 配信 。 - ^ “
鳥 は恐竜 から進化 した ー論争 についに終止符 | Nature ダイジェスト | Nature Portfolio”. www.natureasia.com. 2022年 1月 24日 閲覧 。 - ^ “
驚 きの恐竜 展 を開催 、もはや鳥 展 、米 NYで”. natgeo.nikkeibp.co.jp. 2022年 1月 24日 閲覧 。 - ^ The functional origin of dinosaur bipedalism: cumulative evidence from bipedally inclined reptiles and disinclined mammals(W Scott Persons:2017)
- ^ ヴォード 2008, p. 253
- ^ A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs(Randall B Irmis:2007)
- ^ 「
濱田 隆 『恐竜 学 への誘 い』」『恐竜 学 がわかる。』朝日新聞社 〈AERA MOOK〉、2001年 、[要 ページ番号 ]頁 。ISBN 4022741163。 - ^ Alvarez, L.W.; Alvarez,W., et al. (1980). “Extraterrestrial Cause for the Cretaceous-Tertiary Extinction”. Science 208 (4448): 1095-1108. doi:10.1126/science.208.4448.1095.
- ^ ジェームズ・ローレンス・パウエル『
白 亜紀 に夜 がくる-恐竜 の絶滅 と現代 地質 学 』寺嶋 英志 、瀬戸口 烈 司 (訳 )、青土 社 、2001年 、[要 ページ番号 ]頁 。ISBN 4791759079。 - ^ ムー (
雑誌 )2008年 1月 号 、学研 [出典 無効 ] - ^ Schulte, Peter (2010). Laia Alegret, Ignacio Arenillas, Jose A. Arz, Penny J. Barton, Paul R. Bown, Timothy J. Bralower, Gail L. Christeson, hilippe Claeys, Charles S. Cockell, Gareth S. Collins, Alexander Deutsch, Tamara J. Goldin, Kazuhisa Goto12, Jose M. Grajales-Nishimura, Richard A. F. Grieve, Sean P. S. Gulick, Kirk R. Johnson, Wolfgang Kiessling, Christian Koeberl, David A. Kring, Kenneth G. MacLeod, Takafumi Matsui, Jay Melosh, Alessandro Montanari, Joanna V. Morgan, Clive R. Neal, Douglas J. Nichols, Richard D. Norris, Elisabetta Pierazzo, Greg Ravizza, Mario Rebolledo-Vieyra, Wolf Uwe Reimold, Eric Robin, Tobias Salge, Robert P. Speijer, Arthur R. Sweet, Jaime Urrutia-Fucugauchi, Vivi Vajda, Michael T. Whalen, Pi S. Willumsen. “The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary”. Science 327 (5970): 1214-1218. doi:10.1126/science.1177265.
- ^
恐竜 やアンモナイト等 の絶滅 は「小惑星 衝突 により発生 したすすによる気候 変動 」が原因 だった東北大学 公式 ホームページ - ^
小惑星 が少 しずれていたら…恐竜 生存 ?東北大 など発表 朝日新聞 - ^ “Asteroid that killed dinosaurs also intensified volcanic eruptions - study” (
英語 ). the Guardian (2015年 10月 2日 ). 2020年 12月26日 閲覧 。 - ^
鳥 は「恐竜 の子孫 」ではありません、恐竜 なんです | ギズモード・ジャパン - ^ コルバート, モラレス & ミンコフ 2004, pp. 199–200
- ^ Carpenter, Kenneth (2018-10-19). “Maraapunisaurus fragillimus, N.G. (formerly Amphicoelias fragillimus), a basal Rebbachisaurid from the Morrison Formation (Upper Jurassic) of Colorado” (
英語 ). Geology of the Intermountain West 5: 227–244. doi:10.31711/giw.v5.pp227-244. ISSN 2380-7601. - ^ Paul, Gregory S.; Larramendi, Asier (2023-04-11). “Body mass estimate of Bruhathkayosaurus and other fragmentary sauropod remains suggest the largest land animals were about as big as the greatest whales” (
英語 ). Lethaia 56 (2): 1–11. doi:10.18261/let.56.2.5. ISSN 0024-1164. - ^ “The size of the BYU 9024 animal”. svpow.com (June 16, 2019). 16 April 2022
時 点 のオリジナルよりアーカイブ。June 16, 2019閲覧 。 - ^ Molina-Perez & Larramendi (2020). Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs. New Jersey: Princeton University Press. p. 36
- ^ “2
億 年 前 の大型 恐竜 、本当 は重 くなかった英 米 研究 ”. AFPBB News. AFP. (2012年 6月 7日 ) 2017年 7月 2日 閲覧 。 - ^ コルバート, モラレス & ミンコフ 2004, pp. 191–194
- ^ コルバート, モラレス & ミンコフ 2004, p. 199
- ^ Theoretical aspects of cursorial adaptations in dinosaurs (Walter P Coombs Jr:1978)
- ^ Dinosauromorph research sheds light on dinosaur evolution(CBC NEWS:2015)
- ^ What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs (MT Carrano:1999)
- ^ ヴォード 2008, p. 249
- ^
平山 廉 2007, pp. 66–67 - ^
疋田 努 2002, p. 72 - ^
平山 廉 2007, p. 70 - ^ ,
松岡 &安藤 2009, p. 11 - ^ ,
松岡 &安藤 2009, p. 21 - ^ フジテレビトリビア
普及 委員 会 『トリビアの泉 〜へぇの本 〜 3』講談社 、2003年 。 - ^ Vinther, J., Briggs, D. E. G., Prum, R. O. and Saranathan, V. 2008 The colour of fossil feathers.
- ^ “
恐竜 の色 はどのようにして復元 するの?|恐竜 ・古 生物 Q&A|FPDM:福井 県立 恐竜 博物館 ”. www.dinosaur.pref.fukui.jp. 2022年 1月 22日 閲覧 。 - ^ “Yale Scientists First to Reveal Flamboyant Colors of a Dinosaur’s Feathers” (
英語 ). YaleNews (2010年 2月 4日 ). 2022年 1月 22日 閲覧 。 - ^ a b
鳥類 の起源 参照 - ^ a b
長谷川 政美 『系統 樹 をさかのぼって見 えてくる進化 の歴史 僕 たちの祖先 を探 す15億 年 の旅 』ベレ出版 〈BERET SCIENCE〉、2014年 、102ほか頁 。ISBN 978-4-86064-410-9。 - ^
骨 「年輪 」、草食 哺乳類 にも=恐竜 冷血 説 を否定 -欧州 チーム[リンク切 れ] - ^ Meike Köhler, Nekane Marín-Moratalla, Jordana Xavier, Ronny Aanes (2012
年 7月 19日 ). “Seasonal bone growth and physiology in endotherms shed light on dinosaur physiology”. Nature. Nature Publishing Group. 2017年 7月 2日 閲覧 。 - ^ スウィーテク 2011, pp. 170–171
- ^ a b c “
恐竜 はどうやって巣 を作 るの?卵 はどんなふうに並 べるの?”.福井 県立 恐竜 博物館 . 2023年 11月16日 閲覧 。 - ^ “
恐竜 の繁殖 戦略 と孵化 方法 の多様 性 ”. Dino World. 2023年 11月16日 閲覧 。 - ^ Plot R. (1677) The natural history of Oxford-shire : Being an essay toward the natural history of England. Chap. V. Of formed stones.
- ^
真鍋 真 (2005)「人間 と恐竜 との出会 い」.恐竜 博 2005-恐竜 から鳥 への進化 .図録 - ^ Buckland W. (1824) Notice on the Megalosaurus or great fossil lizard. Stonesfield. Transactions of the Geological Society of London 1(2):390-396.
- ^ Mantell, Gideon A. (1825). “Notice on the Iguanodon, a newly discovered fossil reptile, from the sandstone of Tilgate forest, in Sussex”. Philosophical Transactions of the Royal Society 115: 179–186. doi:10.1098/rstl.1825.0010. ISSN 0261-0523. JSTOR 107739.
- ^ von Meyer, H. (1861). "Archaeopteryx litographica (Vogel-Feder) und Pterodactylus von Solenhofen." Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde: 678-679.
- ^ Huxley, Thomas H. (1868). "On the animals which are most nearly intermediate between birds and reptiles". Annals of the Magazine of Natural History 4 (2): 66–75.
- ^ Heilmann, Gerhard (1926). The Origin of Birds. London: Witherby. pp. 208pp
- ^ Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. JHU Press. ISBN 978-0-8018-6763-7
- ^ Padian, Kevin. (2004). “Basal Avialae”. In Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 978-0-520-24209-8
- ^ Camp, Charles L. (1936). “A new type of small theropod dinosaur from the Navajo Sandstone of Arizona”. Bulletin of the University of California Department of Geological Sciences 24: 39–65.
- ^ Ostrom, John H. (1973). "The ancestry of birds". Nature 242 (5393): 136. Bibcode:1973Natur.242..136O. doi:10.1038/242136a0.
- ^ Gauthier, Jacques. (1986). "Saurischian monophyly and the origin of birds". In Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight. Memoirs of the California Academy of Sciences 8. pp. 1–55.
- ^ Holtz, Jr., T.R. (2000). "Classification and evolution of the dinosaur groups". In Paul, G.S. The Scientific American Book of Dinosaurs. St. Martin's Press. pp. 140–168.
- ^ Burke, A. C. and A. Feduccia (1997). "Developmental patterns and the identification of homologies in the avian hand." Science 278(5338): 666-668.
- ^ Wagner, G. P., and J. Gauthier 1999. 1,2,3 = 2,3,4: a solution to the problem of the homology of the digits in the avian hand. PNAS, 96, 5111-5116.
- ^ Vargas, A. O., T. Kohlsdorf, et al. (2008). "The evolution of HoxD-11 expression in the bird wing: insights from Alligator mississippiensis." PLoS One 3(10): e3325.
- ^ Tamura, K., N. Nomura, et al. (2011). "Embryological evidence identifies wing digits in birds as digits 1, 2, and 3." Science 331(6018): 753-757.
- ^ Xu, X., Clark, J.M., Mo, J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C.-K., Han, F.-L., and Guo, Y. (2009). "A Jurassic ceratosaur from China helps clarify avian digital homologies." Nature, 459(18): 940–944. doi:10.1038/nature08124
- ^ Horner, J. R. and M. B. Goodwin (2008). "Ontogeny of cranial epi-ossifications in Triceratops." Journal of Vertebrate Paleontology 28(1): 134-144.
- ^ Longrich, N. R. and D. J. Field (2012). "Torosaurus is not Triceratops: ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy." PLoS One 7(2): e32623.
- ^ コルバート, モラレス & ミンコフ 2004, p. 190
- ^ MIT’s New Evidence on Dinosaur Evolution(Jennifer Chu:2020)
- ^
疋田 努 2002, p. 43 - ^
金子 隆一 『哺乳類 型 爬虫類 ヒトの知 られざる祖先 』朝日新聞社 〈朝日 選書 〉、1998年 、36頁 。ISBN 4022597097。 - ^ Nagao, Takumi (1936-07). “Nipponosaurus Sachalinensis : A New Genus and Species of Trachodont Dinosaur from Japanese Saghalien”. Journal of the Faculty of Science, Hokkaido Imperial University. Ser. 4, Geology and mineralogy (The Hokkaido Imperial University) 3 (2): 185-220. hdl:2115/35768.
- ^ 『
北海道 中川 町 の恐竜 化石 を新 属 新種 「パラリテリジノサウルス・ジャポニクス」と命名 ~恐竜 類 テリジノサウルス科 の爪 の進化 ~』(プレスリリース)北海道大学 、岡山理科大学 、中川 町 エコミュージアムセンター、2022年 5月 9日 。2022年 5月 9日 閲覧 。 - ^
久保田 克博 「日本 産 の中生代 恐竜 化石 目録 2022年版 」(PDF)『群馬 県立 自然 史 博物館 研究 報告 』第 27号 、群馬 県立 自然 史 博物館 、2023年 3月 、157-170頁 、CRID 1520859470312165120、ISSN 13424092。 - ^
宮原 れい (2019年 9月 6日 ). “通称 ・むかわ竜 、学名 が「カムイサウルス・ジャポニクス」に決定 新 属 新種 の恐竜 だと正式 に認定 される”. ねとらぼ. 2021年 1月 7日 閲覧 。 - ^ Mortimer, Michael (2008
年 ). “Neotheropoda”. The Theropod Database. 2009年 11月17日 閲覧 。 - ^ “
新種 化石 06「桑島 化石 壁 産出 の恐竜 化石 に学名 がつきました」|白山 市 公式 ホームページ”.白山 市 公式 ホームページ. 2023年 3月 22日 閲覧 。 - ^ 「
福井 の恐竜 化石 発掘 調査 30年 きっかけの発見 者 、松田 亜 規 さん」『毎日新聞 』2019年 7月 3日 。2021年 2月 14日 閲覧 。 - ^ Kobayashi, Y. and Azuma, Y. (2003). "A new iguanodontian (Dinosauria; Ornithopoda), form the lower Cretaceous Kitadani Formation of Fukui Prefecture, Japan". Journal of Vertebrate Paleontology 23(1): 166-175
- ^ “フクイラプトルという
学名 になりました”.福井 県立 恐竜 博物館 . 2020年 6月 27日 閲覧 。 - ^ Azuma, Y.; Shibata, M. (2010). “Fukuititan nipponensis, a new titanosauriform sauropod from the Early Cretaceous Tetori Group of Fukui Prefecture, Japan”. Acta Geologica Sinica - English Edition 84 (3): 454–462. doi:10.1111/j.1755-6724.2010.00268.x.
- ^ “
勝山 市 の発掘 現場 から発見 された恐竜 に新 しい学名 が付 けられました”.福井 県立 恐竜 博物館 (2015年 3月 10日 ). 2017年 7月 2日 閲覧 。 - ^ “
新 恐竜 はティラノサウルスもどき 98年 から発掘 の化石 、福井 |全国 のニュース”.福井 新聞 ONLINE. 2023年 9月 8日 閲覧 。 - ^ “
恐竜 博物館 の調査 研究 情報 、長崎 初 の大型 獣 脚 類 (肉食 恐竜 )の化石 について”.福井 県立 恐竜 博物館 (2013年 7月 8日 ). 2017年 7月 2日 閲覧 。 - ^
共同通信 (2019年 1月 28日 ). “国内 最古 級 の恐竜 化石 発見 、熊本 獣 脚 類 の肋骨 か -共同通信 | This Kiji”.共同通信 .共同通信 . 2019年 1月 29日 閲覧 。
参考 文献
- David E. Fastovsky、デヴィッド・ワイシャンペル『
恐竜 学 進化 と絶滅 の謎 』真鍋 真 (訳 )、丸善 、2006年 。ISBN 978-4-621-07734-4。 - David B. Weishampel, Peter Dodson, Halszka Osmólska, ed (2004). The Dinosauria (second edition ed.). University of California Press. ISBN 978-0520242098
- ピーター・D・ヴォード『
恐竜 はなぜ鳥 に進化 したのか』垂水 雄二 (訳 )、文藝春秋 、2008年 。ISBN 978-4-16-369960-8。 - エドウィン・ハリス・コルバート、マイケル・モラレス、イーライ・C・ミンコフ『
脊椎動物 の進化 』田 隅 本 生 (訳 )、築地 書 館 、2004年 。ISBN 4-8067-1295-7。 - ブライアン・スウィーテク『
移行 化石 の発見 』野中 香 方子 (訳 )、文藝春秋 、2011年 。ISBN 978-4-16-373970-0。 疋田 努 『爬虫類 の進化 』東京大学 出版 会 、2002年 。ISBN 4-13-060179-2。平山 廉 『カメのきた道 甲羅 に秘 められた2億 年 の生命 進化 』日本 放送 出版 協会 〈NHKブックス〉、2007年 。ISBN 978-4140910955。松 岡廣 繁 、安藤 みき子 『鳥 の骨 探 』エヌ・ティー・エス、2009年 。ISBN 978-4-86043-276-8。伊地知 英信 『日本 にも恐竜 がいた』長谷川 善和 監修 、どうぶつ社 、1990年 。ISBN 4886224156。
関連 項目
外部 リンク
恐竜 センター (群馬 県 神 流 町 恐竜 センター)恐竜 図鑑 (FPDM福井 県立 恐竜 博物館 )白山 恐竜 パーク白峰 (石川 県 白山 市 )- いわき
市 石炭 化石 館 (福島 県 いわき市 ) 人 と自然 の博物館 (兵庫 県 )恐竜 発掘 project(日本 テレビ系 テレビ番組 『ザ!鉄腕 !DASH!!』)恐竜 渓谷 ふくい勝山 ジオパーク(福井 県 勝山 市 )- ディノ・ネット デジタル
恐竜 展示 室 (国立 科学 博物館 、凸版印刷 ) -博物館 や骨格 等 をオンラインで確認 できるサイト - 『
恐竜 』 - コトバンク