ティラノサウルス科 か (ティラノサウルスか、学名 がくめい Tyrannosauridae ティラノサウルス上 うえ 科 か に属 ぞく 獣 しし 脚 あし 類 るい 恐竜 きょうりゅう 科 か 同上 どうじょう 科 か 恐竜 きょうりゅう 中 なか 大型 おおがた 超 ちょう 大型 おおがた 後期 こうき 白 はく 亜紀 あき 北 きた アメリカ大陸 あめりかたいりく アジア に生息 せいそく 解剖 かいぼう 学 がく 的 てき 特徴 とくちょう 共通 きょうつう 性 せい 高 たか 幼体 ようたい 小型 こがた 属 ぞく 種 しゅ 体 からだ 華奢 きゃしゃ 大型 おおがた 属 ぞく 種 しゅ 成体 せいたい 頑強 がんきょう 体格 たいかく 有 ゆう [ 2]
より小型 こがた 祖先 そせん の子孫 しそん 拘 かか 科 か 恐竜 きょうりゅう 常時 じょうじ 各々 おのおの 生態 せいたい 系 けい 最大 さいだい 捕食 ほしょく 動物 どうぶつ 食物 しょくもつ 連鎖 れんさ 頂点 ちょうてん 位置 いち 最大 さいだい 種 たね Tyrannosaurus rex で、これは既知 きち 陸上 りくじょう 捕食 ほしょく 動物 どうぶつ 最大 さいだい 最 もっと 頑強 がんきょう 最大 さいだい 全長 ぜんちょう 達 たっ [ 3] 最大 さいだい 体重 たいじゅう 推定 すいてい [ 3] [ 4] [ 5] [ 注 ちゅう ] 二 に 足 そく 歩行 ほこう 性 せい 顎 あご 字 じ 型 がた 断面 だんめん 持 も 歯 は 字 じ 型 がた 配列 はいれつ 先端 せんたん 丸 まる 帯 お 長 なが 後肢 あとあし 他 た 大型 おおがた 獣 じゅう 脚 あし 類 るい 比較 ひかく 移動 いどう 速度 そくど 向上 こうじょう 示唆 しさ 一方 いっぽう 前肢 ぜんし 手 て 機能 きのう 保持 ほじ 退化 たいか 指 ゆび 退 すさ 縮 ちぢみ 本 ほん [ 2]
Deinodon の歯 は 最初 さいしょ 期 き 知 し 科 か 化石 かせき ティラノサウルス科 か 最初 さいしょ 化石 かせき 発見 はっけん カナダ地質調査所 ちしつちょうさしょ (英語 えいご 版 ばん 遠征 えんせい 際 さい 際 さい 無数 むすう 離散 りさん 歯 は 発見 はっけん 明 あき 恐竜 きょうりゅう 歯 は 年 ねん ジョセフ・ライディ が Deinodon 怖 こわ 歯 ぱ 命名 めいめい 科 か 最初 さいしょ 良好 りょうこう 標本 ひょうほん アルバータ州 しゅう のホースシューキャニオン累 るい 層 そう で発見 はっけん 部分 ぶぶん 的 てき 骨格 こっかく 伴 ともな 完全 かんぜん 頭蓋骨 ずがいこつ 構成 こうせい 化石 かせき 年 ねん エドワード・ドリンカー・コープ が最初 さいしょ 研究 けんきゅう 東方 とうほう 上 うえ 科 か 属 ぞく ドリプトサウルス の種 たね 考 かんが 年 ねん ヘンリー・フェアフィールド・オズボーン はアルバータ州 しゅう 化石 かせき 顕著 けんちょ 異 こと 認識 にんしき Albertosaurus sarcophagus 肉 にく 食 た 命名 めいめい [ 6] 年 ねん 他 た 類 るい 骨 ほね 記載 きさい 記載 きさい 単 たん 離 はな 椎骨 ついこつ Manospondylus gigas 命名 めいめい 発見 はっけん 年 ねん 以上 いじょう 亘 わた 見落 みお 本 ほん 標本 ひょうほん 実際 じっさい Tyrannosaurus rex 当該 とうがい 学名 がくめい 対 たい 先取 せんしゅ 権 けん 持 も 判明 はんめい 年代 ねんだい 初頭 しょとう 議論 ぎろん 呼 よ [ 7]
アウブリソドン に割 わ 当 あ 歯 は 1905年 ねん 命名 めいめい 論文 ろんぶん 発表 はっぴょう 同年 どうねん 類 るい 標本 ひょうほん 記載 きさい 標本 ひょうほん アメリカ自然 しぜん 史 し 博物館 はくぶつかん が1902年 ねん 行 い バーナム・ブラウン 率 ひき 遠征 えんせい モンタナ州 しゅう とワイオミング州 しゅう で発見 はっけん 当初 とうしょ 標本 ひょうほん 別種 べっしゅ 考 かんが 彼 かれ 標本 ひょうほん Dynamosaurus imperiosus (「力 ちから 皇帝 こうてい 標本 ひょうほん Tyrannosaurus rex (「暴君 ぼうくん 王 おう 命名 めいめい 年 ねん 後 ご 標本 ひょうほん 同 おな 種 しゅ 由来 ゆらい 認識 にんしき 最初 さいしょ 発見 はっけん 方 ほう 名前 なまえ 原 げん 記載 きさい 先 さき 記載 きさい 名前 なまえ 用 もち [ 8]
バーナム・ブラウンはアルバータ州 しゅう 科 か 標本 ひょうほん 蒐集 しゅうしゅう 当該 とうがい 特徴 とくちょう 的 てき 本 ほん 指 ゆび 短 みじか 前肢 ぜんし 保存 ほぞん 年 ねん ローレンス・ラム が Gorgosaurus libratus 均衡 きんこう 熾烈 しれつ 命名 めいめい 中 なか 含 ふく 関 かか 目 め 重大 じゅうだい 発見 はっけん 年 ねん 保存 ほぞん 状態 じょうたい 良好 りょうこう 異様 いよう 小型 こがた 完全 かんぜん 頭蓋骨 ずがいこつ 発見 はっけん 当該 とうがい 標本 ひょうほん 第 だい 二 に 次 じ 世界 せかい 大戦 たいせん 後 のち チャールズ・W・ギルモア が研究 けんきゅう Gorgosaurus lancesnis と命名 めいめい [ 6] 年 ねん ロバート・T・バッカー とフィリップ・J・カリー およびマイケル・ウィリアムズがこの頭蓋骨 ずがいこつ 再 さい 研究 けんきゅう 新 しん 属 ぞく Nanotyrannus 分類 ぶんるい [ 9] 年 ねん ソビエト連邦 れんぽう の古 こ 生物 せいぶつ 学者 がくしゃ モンゴル国 こく への遠征 えんせい 開始 かいし 初 はつ 類 るい 化石 かせき 発見 はっけん 年 ねん 化石 かせき 新 しん 属 ぞく Tarbosaurus 怖 こわ 命名 めいめい 後 ご 研究 けんきゅう 類 るい 種 たね 実際 じっさい 成長 せいちょう 段階 だんかい 異 こと 種 たね 判明 はんめい 後 のち 発見 はっけん 第 だい 二 に 種 たね 年 ねん 記載 きさい 命名 めいめい Alioramus remotus 隔 へだ 異 こと 枝 えだ 真 しん 科 か 基盤 きばん 的 てき 類 るい 議論 ぎろん [ 10] [ 6]
ティラノサウルス 、タルボサウルス 、アルバートサウルス 、ゴルゴサウルス 、ダスプレトサウルス の大 おお 比較 ひかく ティラノサウルス科 か 大型 おおがた 動物 どうぶつ 全 すべ 種 たね 体重 たいじゅう 超 こ [ 11] アリオラムス の1個 いっこ 標本 ひょうほん 全長 ぜんちょう 個体 こたい 推定 すいてい 発見 はっけん [ 10] 未 み 成熟 せいじゅく 個体 こたい 考 かんが 専門 せんもん 家 か [ 11] [ 12] アルバートサウルス 、ゴルゴサウルス 、ダスプレトサウルス はいずれも全長 ぜんちょう [ 13] タルボサウルス は10メートル以上 いじょう 達 たっ [ 14] 最大 さいだい 級 きゅう 標本 ひょうほん 一 ひと スー は全長 ぜんちょう 達 たっ [ 3]
部分 ぶぶん 的 てき 頭蓋骨 ずがいこつ 発見 はっけん 除 のぞ 全 すべ 属 ぞく 完全 かんぜん 頭蓋骨 ずがいこつ 得 え 科 か 頭蓋骨 ずがいこつ 解剖 かいぼう 学 がく 理解 りかい [ 15] 頭蓋骨 ずがいこつ 長 なが 超 こ [ 13] 成体 せいたい 科 か 高 たか 頑強 がんきょう 頭蓋骨 ずがいこつ 持 も 数 すう 多 おお 骨 ほね 癒合 ゆごう 強度 きょうど 増 ま 同時 どうじ 数 すう 多 おお 頭蓋骨 ずがいこつ 間 あいだ 存在 そんざい 空洞 くうどう 骨 ほね 間 あいだ 位置 いち 大型 おおがた 開口 かいこう 部 ぶ 窓 まど 頭蓋骨 ずがいこつ 軽量 けいりょう 化 か 寄与 きよ 高 たか 前 ぜん 上顎 じょうがく 骨 こつ 癒合 ゆごう 鼻骨 びこつ 科 か 頭蓋骨 ずがいこつ 見 み 数 すう 多 おお 特徴 とくちょう 直近 ちょっきん 祖先 そせん 見 み 出来 でき [ 11]
ティラノサウルス科 か 頭蓋骨 ずがいこつ 数 すう 多 おお 固有 こゆう 形質 けいしつ 具体 ぐたい 的 てき 卓越 たくえつ 矢 や 状 じょう 稜 りょう (英語 えいご 版 ばん 伴 ともな 癒合 ゆごう 頭頂 とうちょう 骨 こつ 挙 あ 稜 りょう 矢 や 状 じょう 縫合 ほうごう (英語 えいご 版 ばん 沿 そ 縦 たて 方向 ほうこう 走 はし 頭蓋 とうがい 天井 てんじょう 上 じょう 側 がわ 頭 あたま 窓 まど 隔 へだ 窓 まど 後 のち 側 がわ 科 か 特徴 とくちょう 的 てき 高 たか 項 こう 部 ぶ 持 も 縦 たて 方向 ほうこう 横断 おうだん 面 めん 沿 そ 頭頂 とうちょう 骨 こつ 上 じょう 走 はし 発達 はったつ 涙 なみだ 骨 こつ 上 うえ 目 め 前 まえ 高 たか 持 も 極端 きょくたん 厚 あつ 帯 お 後 こう 眼窩 がんか 骨 こつ 三日月 みかづき 形 がた 目 め 後 うし 形成 けいせい 最 さい 上部 じょうぶ 列 れつ 配列 はいれつ 個 こ 骨質 こっしつ 持 も 低 ひく 報告 ほうこく 標本 ひょうほん 基盤 きばん 的 てき 上 うえ 科 か アパラチオサウルス とも共通 きょうつう [ 12] [ 16] 頭蓋骨 ずがいこつ 他 ほか 部位 ぶい 無数 むすう 孔 あな 存在 そんざい D. horneri 記載 きさい 年 ねん 研究 けんきゅう ワニ やティラノサウルス科 か 見 み 多 た 列 れつ 神経 しんけい 血管 けっかん 孔 あな 触覚 しょっかく 鱗状 りんじょう 外皮 がいひ 相関 そうかん [ 17]
ティラノサウルス科 か 上 うえ 科 か 祖先 そせん 同様 どうよう 異 い 歯 は 性 せい 持 も 前上 まえかみ 顎骨 がっこつ 歯 ぱ 断面 だんめん 字 じ 型 がた 他 た 歯 は 小型 こがた 最初 さいしょ 期 き 上 うえ 科 か 他 た 獣 しし 脚 あし 類 るい 異 こと 成熟 せいじゅく 科 か 上顎 じょうがく 骨 こつ 歯 は 歯 は 骨 こつ 歯 は 状 じょう 極端 きょくたん 厚 あつ 断面 だんめん 丸 まる 帯 お 種 たね 鋸歯 きょし 減少 げんしょう [ 11] 歯 は 本数 ほんすう 種 たね 一定 いってい 大型 おおがた 種 たね 小型 こがた 種 たね 少 すく 傾向 けいこう 例 たと 本 ほん 本 ほん 歯 は 持 も [ 18]
ウィリアム・エイブラーは2001年 ねん 歯 は 鋸歯 きょし 歯 は 割 わ 目 め 類似 るいじ 膨大 ぼうだい 部 ぶ 呼称 こしょう 丸 まる 空洞 くうどう 帰着 きちゃく 観察 かんさつ [ 19] 科 か 歯 は 体 からだ 肉 にく 引 ひ 張 ぱ 留 と 具 ぐ 用 もち 類 るい 肉片 にくへん 引 ひ 剥 は 際 さい 張力 ちょうりょく 割 わ 目 め 鋸歯 きょし 歯 は 一 いち 面 めん 広 ひろ 可能 かのう 性 せい [ 19] 膨大 ぼうだい 部 ぶ 力 ちから 大 おお 表面積 ひょうめんせき 分散 ぶんさん 負担 ふたん 歯 は 損傷 そんしょう 軽減 けいげん [ 19] 空洞 くうどう 帰着 きちゃく 切 き 込 こ 存在 そんざい 人間 にんげん 工学 こうがく 相 そう 等 ひと 説明 せつめい ギター メーカーは空洞 くうどう 帰着 きちゃく 切 き 込 こ 入 い 柔軟 じゅうなん 性 せい 剛性 ごうせい 交互 こうご 木材 もくざい 与 あた [ 19] 航空機 こうくうき 表面 ひょうめん 保護 ほご 膨大 ぼうだい 部 ぶ 様 さま 構造 こうぞう 作 つく 素材 そざい 亀裂 きれつ 伝播 でんぱ 防 ふせ 方法 ほうほう 用 もち [ 19] 切 き 込 こ 穿孔 せんこう 伴 ともな アクリル樹脂 じゅし のバーが規則 きそく 的 てき 切 き 込 こ 入 い 比 くら 以上 いじょう 強 つよ 強度 きょうど 持 も 実証 じっしょう [ 19] 類 るい 他 た 獣 しし 脚 あし 類 るい 異 こと 植 うえ 竜 りゅう 類 るい ディメトロドン などの太古 たいこ 捕食 ほしょく 動物 どうぶつ 摂食 せっしょく 力 ちから 受 う 歯 は 切 き 目 め 状 じょう 鋸歯 きょし 広 ひろ 適応 てきおう 遂 と [ 19]
リトロナクス とテラトフォネウス の骨格 こっかく 頭蓋骨 ずがいこつ 厚 あつ 字 じ 型 がた 先端 せんたん 位置 いち 長 なが 重厚 じゅうこう 尾 お 頭部 とうぶ 胴 どう 部 ぶ 対 たい カウンターウエイト として機能 きのう 重心 じゅうしん 臀部 でんぶ 位置 いち 科 か 的 てき 非常 ひじょう 小型 こがた 本 ほん 指 ゆび 前肢 ぜんし 知 し 痕跡 こんせき 器官 きかん 第 だい 指 ゆび 残 のこ 場合 ばあい [ 11] [ 20]
ゴルゴサウルス の肩 かた 甲 かぶと 骨 こつ 右 みぎ 前肢 ぜんし ティラノサウルス科 か 後肢 あとあし 歩行 ほこう 骨 ほね 頑強 がんきょう 前肢 ぜんし 対照 たいしょう 的 てき 大半 たいはん 他 ほか 獣 しし 脚 あし 類 るい 比較 ひかく 後肢 あとあし 体 からだ 対 たい 長 なが 幼体 ようたい 小型 こがた 成体 せいたい 基盤 きばん 的 てき 上 うえ 科 か 同様 どうよう 脛骨 けいこつ 大腿 だいたい 骨 こつ 長 なが オルニトミモサウルス類 るい と同様 どうよう 脚 あし 速 はや 恐竜 きょうりゅう 形質 けいしつ 示 しめ 大型 おおがた 成体 せいたい 動作 どうさ 緩慢 かんまん 動物 どうぶつ 後肢 あとあし 形質 けいしつ 有 ゆう アベリサウルス科 か やカルノサウルス類 るい のような他 ほか 大型 おおがた 獣 じゅう 脚 あし 類 るい 見 み 科 か 第 だい 中 ちゅう 足 あし 骨 こつ 第 だい 第 だい 中 ちゅう 足 あし 骨 こつ 挟 はさ 細 ぼそ アークトメタターサル として知 し 構造 こうぞう [ 11]
アークトメタターサルが最初 さいしょ 進化 しんか 時代 じだい 不明 ふめい ディロング のような最初 さいしょ 期 き 上 うえ 科 か 存在 そんざい [ 21] 新 あたら 時代 じだい アパラチオサウルス には見 み [ 16] 構造 こうぞう トロオドン科 か やオルニトミモサウルス類 るい およびカエグナトゥス科 か (英語 えいご 版 ばん 特徴 とくちょう [ 22] 最初 さいしょ 期 き 上 うえ 科 か 存在 そんざい 当該 とうがい 形質 けいしつ 収斂 しゅうれん 進化 しんか 示唆 しさ [ 21]
Deinodontidae という名前 なまえ 年 ねん 科 か 作 つく [ 23] 年代 ねんだい 新 あら 名前 なまえ 立 た 位置 いち 用 もち [ 24] 属 ぞく Deinodon モンタナ州 しゅう 産 さん 単 たん 離 はな 歯 は 命名 めいめい [ 25] 年 ねん 北 きた 類 るい デイル・ラッセル は Deinodon が有効 ゆうこう 分類 ぶんるい 群 ぐん 結論 けつろん 代 か 名前 なまえ 用 もち 国際 こくさい 動物 どうぶつ 命名 めいめい 規約 きやく 一致 いっち 主張 しゅちょう [ 13] 科 か 名前 なまえ 現在 げんざい 専門 せんもん 家 か 好 この [ 6]
Tyrannosaurus は1905年 ねん ヘンリー・フェアフィールド・オズボーン が命名 めいめい 伴 ともな 科 か 命名 めいめい [ 26] 名前 なまえ 古代 こだい 語 ご τυραννος (tyrannos 暴君 ぼうくん σαυρος (sauros 由来 ゆらい 接尾 せつび 詞 し -idae は動物 どうぶつ 科 か 名前 なまえ 一般 いっぱん 付 ふ 複数 ふくすう 名詞 めいし 示 しめ 語 ご 接尾 せつび 詞 し -ιδαι -idai に由来 ゆらい [ 27]
一般 いっぱん 化 か 差異 さい 示 しめ ティラノサウルス科 か リンネ式 しき 階層 かいそう 分類 ぶんるい 体系 たいけい における科 か ティラノサウルス上 うえ 科 か と獣 しし 脚 あし 亜 あ 目 め 内部 ないぶ 置 お
ティラノサウルス科 か 亜 あ 科 か 細分 さいぶん アルバートサウルス亜 あ 科 か は北 きた 属 ぞく アルバートサウルス とゴルゴサウルス を含 ふく ティラノサウルス亜 あ 科 か はダスプレトサウルス 、テラトフォネウス 、ビスタヒエヴェルソル 、タルボサウルス 、ナヌークサウルス 、ズケンティラヌス 、ティラノサウルス を含 ふく [ 28] Gorgosaurus libratus をアルバートサウルス属 ぞく Tarbosaurus bataar をティラノサウルス属 ぞく 含 ふく 研究 けんきゅう 者 しゃ [ 16] [ 6] [ 29] 分 わ 属 ぞく 維持 いじ 好 この 研究 けんきゅう 者 しゃ [ 11] [ 12] 亜 あ 科 か 亜 あ 科 か 比較 ひかく 細身 ほそみ 体格 たいかく 低 ひく 頭蓋骨 ずがいこつ 的 てき 長 なが 脛骨 けいこつ 特徴 とくちょう 持 も [ 11] 亜 あ 科 か 頭頂 とうちょう 骨 こつ 矢 や 状 じょう 前 ぜん 頭骨 とうこつ 上 うえ 続 つづ [ 12] 年 ねん et al. がティラノサウルス科 か 内 ない 族 ぞく アリオラムス とキアンゾウサウルス を含 ふく 族 ぞく 記載 きさい 彼 かれ 系統 けいとう 解析 かいせき 本 ほん 族 ぞく 亜 あ 科 か 基底 きてい 置 お 示唆 しさ [ 30] [ 31] 他 ほか 研究 けんきゅう 者 しゃ 様々 さまざま 科 か 恐竜 きょうりゅう 組 く 合 あ 亜 あ 科 か 他 ほか 下位 かい 分類 ぶんるい 族 ぞく 作成 さくせい [ 32] [ 33] 系統 けいとう 学 がく 的 てき 定義 ていぎ 構成 こうせい 属 ぞく 他 た 属 ぞく 種 たね 考 かんが [ 18]
さらなる亜 あ 科 か 断片 だんぺん 的 てき 属 ぞく 命名 めいめい アウブリソドン 亜 あ 科 か デイノドン 亜 あ 科 か 挙 あ 通例 つうれい 疑問 ぎもん 名 めい 考 かんが 彼 かれ 彼 かれ 亜 あ 科 か 科 か 分類 ぶんるい 通常 つうじょう 除外 じょがい 科 か 当初 とうしょ 基盤 きばん 的 てき 上 うえ 科 か 考 かんが 類似 るいじ 亜 あ 科 か 幼体 ようたい 代表 だいひょう 可能 かのう 性 せい 高 たか ラプトレックス がいる。しかし、本属 ほんぞく 幼体 ようたい 標本 ひょうほん 知 し 疑問 ぎもん 名 めい 考 かんが [ 34]
古 こ 脊椎動物 せきついどうぶつ 学 がく 系統 けいとう 分類 ぶんるい 学 がく 出現 しゅつげん 科 か 複数 ふくすう 明白 めいはく 定義 ていぎ 与 あた 最初 さいしょ 定義 ていぎ 年 ねん アレクトロサウルス やアウブリソドン あるいはナノティラヌス よりもティラノサウルスに近 きん 縁 えん 全 すべ 上 うえ 科 か 本科 ほんか 含 ふく [ 35] 幼体 ようたい 考 かんが 疑問 ぎもん 名 めい 系統 けいとう 群 ぐん 定義 ていぎ 不適 ふてき [ 11]
2001年 ねん 科 か 分岐 ぶんき 分析 ぶんせき 発表 はっぴょう [ 36] 彼 かれ 鋸歯 きょし 無 な 前 ぜん 上顎 じょうがく 骨 こつ 歯 は 特徴 とくちょう 原始 げんし 的 てき 亜 あ 科 か 亜 あ 科 か 亜 あ 科 か 存在 そんざい 結論 けつろん [ 36] 亜 あ 科 か 他 ほか Kirtland Aublysodon " とアレクトロサウルス が含 ふく [ 36] シャモティラヌス がティラノサウルス科 か 共有 きょうゆう 派生 はせい 形質 けいしつ 複数 ふくすう 示 しめ 発見 はっけん 科 か 置 お [ 36]
同誌 どうし 後述 こうじゅつ 記述 きじゅつ 彼 かれ 科 か 最 もっと 近 ちか 共通 きょうつう 祖先 そせん 全 すべ 子孫 しそん 定義 ていぎ 提唱 ていしょう [ 36] 彼 かれ 他 た 研究 けんきゅう 者 しゃ 過去 かこ 提唱 ていしょう 定義 ていぎ 批判 ひはん [ 36] 幼体 ようたい 誤 あやま 同定 どうてい 考 かんが 提唱 ていしょう 定義 ていぎ 科 か 属 ぞく 下位 かい 分類 ぶんるい [ 36] 提案 ていあん 定義 ていぎ 亜 あ 科 か 限定 げんてい [ 36]
2003年 ねん アルバートサウルス 、アレクトロサウルス 、アリオラムス 、ダスプレトサウルス 、ゴルゴサウルス 、タルボサウルス およびティラノサウルスを定義 ていぎ 加 くわ [ 37] 年 ねん 系統 けいとう 群 ぐん 再 さい 定義 ていぎ 彼 かれ 分析 ぶんせき 確固 かっこ 位置 いち 置 お 除 のぞ 上記 じょうき 全 すべ 指示 しじ 子 こ 用 もち 彼 かれ 同 おな 論文 ろんぶん エオティラヌス よりもティラノサウルスに近 きん 縁 えん 全 すべ 獣 しし 脚 あし 類 るい 含 ふく 別 べつ 定義 ていぎ 提供 ていきょう [ 11] 年 ねん 定義 ていぎ 科 か 含 ふく 最 もっと 包括 ほうかつ 的 てき 系統 けいとう 群 ぐん 定義 ていぎ [ 38]
ティラノサウルス科 か 系統 けいとう 分岐 ぶんき 分析 ぶんせき 姉妹 しまい 群 ぐん 置 お 基盤 きばん 的 てき 位置 いち 置 お 近 きん 縁 えん 性 せい 多数 たすう 頭蓋骨 ずがいこつ 特徴 とくちょう 支持 しじ 特定 とくてい 骨 ほね 縫合 ほうごう 線 せん 目 め 後 うし 後 のち 眼窩 がんか 骨 こつ 三日月 みかづき 型 がた 存在 そんざい 下端 かたん 下 した 側 がわ 顕著 けんちょ 湾曲 わんきょく 非常 ひじょう 深 ふか 上顎 じょうがく 骨 こつ [ 11] [ 16] 年 ねん 提唱 ていしょう 別 べつ 仮説 かせつ 鼻骨 びこつ 涙 なみだ 骨 こつ 接続 せつぞく 骨質 こっしつ 先端 せんたん 部 ぶ 存在 そんざい 基 もと 産 さん 含 ふく 系統 けいとう 群 ぐん 基盤 きばん 的 てき 弱 よわ 支持 しじ 示 しめ [ 18]
関連 かんれん 研究 けんきゅう 属 ぞく 間 あいだ 共有 きょうゆう 下 した 顎 あご 固定 こてい 機 き 序 じょ 指摘 してき [ 39] 別 べつ 論文 ろんぶん 幼体 ようたい 可能 かのう 性 せい 指摘 してき 卓越 たくえつ 鼻骨 びこつ 本数 ほんすう 多 おお 歯 は 別 べつ 属 ぞく 示唆 しさ 主張 しゅちょう 同様 どうよう 歯 は 本数 ほんすう 用 もち 同属 どうぞく 別 べつ 属 ぞく 可能 かのう 性 せい 示唆 しさ [ 12] キアンゾウサウルス の発見 はっけん 記載 きさい 近 きん 縁 えん 新 あら 記載 きさい 族 ぞく 属 ぞく 存在 そんざい 長 なが 科 か 恐竜 きょうりゅう 幅広 はばひろ 分布 ぶんぷ 異 こと 獲物 えもの 狩 か 大型 おおがた 頑強 がんきょう 亜 あ 科 か 競合 きょうごう 回避 かいひ 環境 かんきょう 共有 きょうゆう 明 あき [ 40]
Body mass (kg)
6,000
5,000
4,000
3,000
2,000
1,000
5
10
15
20
25
30
Age (years)
Hypothesized growth curves (body mass versus age) of four tyrannosaurids
[ 44]
古 こ 生物 せいぶつ 学者 がくしゃ 科 か 成長 せいちょう 生活 せいかつ 史 し 研究 けんきゅう 分析 ぶんせき 標本 ひょうほん 死亡 しぼう 際 さい 年齢 ねんれい 決定 けってい 様々 さまざま 個体 こたい 年齢 ねんれい 彼 かれ 大 おお 対 たい 上 じょう 成長 せいちょう 率 りつ 調査 ちょうさ 長 なが 幼体 ようたい 期間 きかん 過 す 後 のち 一 いち 生涯 しょうがい 途中 とちゅう 約 やく 年間 ねんかん 驚異 きょうい 的 てき 成長 せいちょう 期 き 迎 むか 明 あき 性 せい 成熟 せいじゅく 急 きゅう 成長 せいちょう 期 き 終 お 成体 せいたい 成長 せいちょう 鈍化 どんか 類 るい 成長 せいちょう 曲線 きょくせん 字 じ 型 がた 歳 さい 前後 ぜんこう 個体 こたい 成長 せいちょう 速度 そくど 最大 さいだい [ 44]
ジェーン の骨格 こっかく 既知 きち 個体 こたい 最小 さいしょう 体重 たいじゅう 僅 わず 歳 さい 個体 こたい 一方 いっぽう 最大 さいだい 級 きゅう 個体 こたい FMNH PR2081(スー )であり、体重 たいじゅう 約 やく 年齢 ねんれい 歳 さい 推定 すいてい 本 ほん 種 しゅ 上限 じょうげん 近 ちか 可能 かのう 性 せい [ 44] 幼体 ようたい 約 やく 歳 さい 体重 たいじゅう 未満 みまん 約 やく 歳 さい 劇的 げきてき 体 からだ 増大 ぞうだい 急速 きゅうそく 成長 せいちょう 期 き 間 あいだ 若 わか 年間 ねんかん 年間 ねんかん 平均 へいきん 体重 たいじゅう 増加 ぞうか 経験 けいけん 成長 せいちょう 速度 そくど 歳 さい 低下 ていか 歳 さい 再 ふたた 達 たっ 劇的 げきてき 成長 せいちょう 減速 げんそく [ 45] 例 たと 歳 さい 歳 さい 標本 ひょうほん RTMP 81.12.1)の間 あいだ 僅 わず 差異 さい [ 44] 急激 きゅうげき 成長 せいちょう 速度 そくど 変化 へんか 肉体 にくたい 的 てき 成熟 せいじゅく 示唆 しさ 可能 かのう 性 せい 仮説 かせつ モンタナ州 しゅう 産 さん 歳 さい 標本 ひょうほん Bレックス )の大腿 だいたい 骨 こつ 骨髄 こつづい 組織 そしき 発見 はっけん 支持 しじ [ 46] 骨髄 こつづい 組織 そしき 排卵 はいらん 期 き 雌 めす 鳥類 ちょうるい 確認 かくにん 繁殖 はんしょく 期 き 示唆 しさ [ 47]
他 た 酷似 こくじ 成長 せいちょう 曲線 きょくせん 示 しめ 成体 せいたい 比較 ひかく 小 ちい 成長 せいちょう 速度 そくど 対応 たいおう [ 48] 亜 あ 科 か 比較 ひかく 急速 きゅうそく 成長 せいちょう 期 き 成長 せいちょう 速度 そくど 高 たか 成体 せいたい 体重 たいじゅう 重 おも 成体 せいたい 体重 たいじゅう 推定 すいてい 基 もと 最大 さいだい 成長 せいちょう 速度 そくど 年間 ねんかん 他 た 研究 けんきゅう 者 しゃ 体重 たいじゅう 重 おも 見積 みつ 成長 せいちょう 速度 そくど 規模 きぼ 左右 さゆう 全体 ぜんたい 変化 へんか 考 かんが [ 44] 最 もっと 若 わか 既知 きち 発見 はっけん 歳 さい 個体 こたい 体重 たいじゅう 約 やく 推定 すいてい 全長 ぜんちょう 僅 わず 上回 うわまわ 同 おな 産地 さんち 既知 きち 最大 さいだい 最 さい 高齢 こうれい 個体 こたい 発見 はっけん 全長 ぜんちょう 年齢 ねんれい 歳 さい 最速 さいそく 成長 せいちょう 速度 そくど 歳 さい 迎 むか 推定 すいてい 体重 たいじゅう 成体 せいたい 基 もと 速度 そくど 年間 ねんかん 達 たっ 見積 みつも 成長 せいちょう 速度 そくど 五 ご 分 ぶん 一 いち 計算 けいさん 最大 さいだい 成長 せいちょう 速度 そくど 急 きゅう 成長 せいちょう 段階 だんかい 約 やく 匹敵 ひってき [ 44]
いまだ未 み 確認 かくにん 属 ぞく 類 るい 胚 はい 発見 はっけん 科 か 卵 たまご 中 なか 発生 はっせい 段階 だんかい 特徴 とくちょう 的 てき 骨格 こっかく 構造 こうぞう 発達 はったつ 示唆 しさ 年 ねん 州 しゅう 層 そう 発見 はっけん 下 しも 顎 あご 歯 は 年 ねん 層 そう 発見 はっけん 年 ねん 記載 きさい 足 あし 鉤 かぎ 爪 つめ 標本 ひょうほん 大 おお 科 か 新生児 しんせいじ 近 ちか 大 おお 齧 かじ 歯 は 類 るい 全身 ぜんしん 同等 どうとう 規模 きぼ 頭蓋骨 ずがいこつ 伴 ともな 生 う 全身 ぜんしん 小型 こがた 犬 いぬ 程度 ていど 大 おお 可能 かのう 性 せい 考 かんが 下 した 顎 あご 標本 ひょうほん 全長 ぜんちょう 約 やく 個体 こたい 鉤 かぎ 爪 つめ 全長 ぜんちょう 約 やく 個体 こたい 考 かんが 標本 ひょうほん 関連 かんれん 卵殻 らんかく 未 み 発見 はっけん 科 か 新生児 しんせいじ 発見 はっけん 場所 ばしょ 動物 どうぶつ 共存 きょうぞん 捕食 ほしょく 他 ほか 種 たね 同 おな 巣 す 利用 りよう 示唆 しさ [ 49] 卵殻 らんかく 標本 ひょうほん 無 な 科 か ムササウルス 属 ぞく プロトケラトプス 属 ぞく 考 かんが 卵 たまご 産 う 可能 かのう 性 せい 指摘 してき [ 50]
ワピチ層 そう 足跡 あしあと 化石 かせき 科 か 爪先 つまさき 厚 あつ 足 あし 幅広 はばひろ 体重 たいじゅう 支持 しじ 成長 せいちょう 示唆 しさ 幅広 はばひろ 足 あし 成体 せいたい 科 か 子孫 しそん 緩慢 かんまん 動 うご 示唆 しさ [ 51] [ 52]
ゴルゴサウルス・リブラトゥスのほぼ完全 かんぜん 骨格 こっかく ロイヤル・ティレル古 こ 生物 せいぶつ 学 がく 博物館 はくぶつかん 急 きゅう 成長 せいちょう 段階 だんかい 終 お 性 せい 成熟 せいじゅく 期 き 入 はい 示唆 しさ 後 ご 一 いち 生涯 しょうがい 通 つう 緩慢 かんまん 成長 せいちょう 続 つづ 示唆 しさ [ 44] [ 48] 活発 かっぱつ 成長 せいちょう 時期 じき 性的 せいてき 成熟 せいじゅく 小型 こがた [ 53] 大型 おおがた [ 54] 恐竜 きょうりゅう ゾウ といった大型 おおがた 哺乳類 ほにゅうるい 共通 きょうつう 形質 けいしつ [ 54] 相対 そうたい 的 てき 性 せい 成熟 せいじゅく 早 はや 成長 せいちょう 完了 かんりょう 後 のち 性 せい 成熟 せいじゅく 遅延 ちえん 鳥類 ちょうるい 顕著 けんちょ 異 こと [ 54] [ 55]
各 かく 年齢 ねんれい 層 そう 標本 ひょうほん 数 すう 集計 しゅうけい 科 か 個体 こたい 群 ぐん 生活 せいかつ 史 し 結論 けつろん 導 みちび 結果 けっか 化石 かせき 記録 きろく 幼体 ようたい 稀 まれ 一方 いっぽう 急 きゅう 成長 せいちょう 期 き 亜 あ 成体 せいたい 成体 せいたい 遥 はる 多 おお 判明 はんめい 半数 はんすう 上回 うわまわ 標本 ひょうほん 性 せい 成熟 せいじゅく 後 ご 年 ねん 以内 いない 死亡 しぼう 他 た 類 るい 現代 げんだい 大型 おおがた 長寿 ちょうじゅ 鳥類 ちょうるい 哺乳類 ほにゅうるい 見 み 種 たね 新生児 しんせいじ 時期 じき 死亡 しぼう 率 りつ 高 たか 次 つ 幼体 ようたい 時期 じき 死亡 しぼう 率 りつ 比較的 ひかくてき 低 ひく 特徴 とくちょう 性 せい 成熟 せいじゅく 後 ご 死亡 しぼう 率 りつ 再 ふたた 上昇 じょうしょう 生殖 せいしょく 一因 いちいん 保存 ほぞん 状態 じょうたい 採集 さいしゅう 起因 きいん 可能 かのう 性 せい 現代 げんだい 大型 おおがた 哺乳類 ほにゅうるい 見 み 一定 いってい 以上 いじょう 大 おお 幼体 ようたい 死亡 しぼう 率 りつ 低 ひく 原因 げんいん 推論 すいろん 歳 さい 同 どう 時代 じだい 全 すべ 捕食 ほしょく 者 しゃ 上回 うわまわ 大 おお 捕食 ほしょく 者 しゃ 不在 ふざい 起因 きいん 死亡 しぼう 率 りつ 低下 ていか 可能 かのう 性 せい 古 こ 生物 せいぶつ 学者 がくしゃ 十分 じゅうぶん 数 かず 化石 かせき 発見 はっけん 同様 どうよう 分析 ぶんせき 不可能 ふかのう 同様 どうよう 一般 いっぱん 的 てき 傾向 けいこう 指摘 してき [ 48]
ティラノサウルス科 か 生涯 しょうがい 半分 はんぶん 幼体 ようたい 過 す 後 のち 数 すう 年 ねん 最大 さいだい 急 きゅう 成長 せいちょう 遂 と [ 44] 巨大 きょだい 成体 せいたい 科 か 他 た 小型 こがた 獣 じゅう 脚 あし 類 るい 中 なか 間 あいだ 的 てき 大 おお 捕食 ほしょく 者 しゃ 完全 かんぜん 欠落 けつらく 生態 せいたい 的 てき 地位 ちい 幼体 ようたい 科 か 埋 う 示唆 しさ 孵化 ふか 後 のち 樹 き 上 じょう 生活 せいかつ 送 おく 緩慢 かんまん 成熟 せいじゅく 大型 おおがた 脊椎動物 せきついどうぶつ 引 ひ 倒 たお 出来 でき 頑丈 がんじょう 頂点 ちょうてん 捕食 ほしょく 者 しゃ 至 いた 現生 げんなま コモドオオトカゲ にも見 み [ 11] 例 たと 群集 ぐんしゅう 発見 はっけん 群 む 異 こと 年齢 ねんれい 個体 こたい 入 はい 混 ま [ 56] [ 57]
移動 いどう 能力 のうりょく 最 もっと 研究 けんきゅう 関 かん 話題 わだい 旋回 せんかい 得意 とくい 直線 ちょくせん 運動 うんどう 最 さい 高速度 こうそくど 年 ねん 研究 けんきゅう 旋回 せんかい 緩慢 かんまん 可能 かのう 性 せい 指摘 してき 度 ど 回転 かいてん 秒 びょう 要 よう 考 かんが 度 ど 直立 ちょくりつ 尾 お 人間 にんげん 秒 びょう 回転 かいてん 可能 かのう 量 りょう [ 58] 旋回 せんかい 苦手 にがて 原因 げんいん 回転 かいてん 慣性 かんせい 質量 しつりょう 多 おお 人間 にんげん 重 おも 材木 ざいもく 運 はこ 重心 じゅうしん 程度 ていど 離 はな 存在 そんざい [ 59] 年 ねん 研究 けんきゅう 違 ちが 見解 けんかい 示 しめ 科 か 他 た 大型 おおがた 獣 じゅう 脚 あし 類 るい ケラトサウルス やカルノサウルス類 るい など)より2倍 ばい 敏捷 びんしょう 小型 こがた 幼体 ようたい 同士 どうし 敏捷 びんしょう 性 せい 差 さ 倍 ばい 指摘 してき 各種 かくしゅ 獣 じゅう 脚 あし 類 るい 腰 こし 脚 あし 部 ぶ 筋肉 きんにく 量 りょう 慣性 かんせい 計算 けいさん 式 しき 推測 すいそく [ 60]
研究 けんきゅう 者 しゃ 最大 さいだい 速度 そくど 推定 すいてい 非常 ひじょう 値 ね 幅 はば 広 ひろ 主 おも 約 やく 毎秒 まいびょう 推定 すいてい 毎秒 まいびょう 毎秒 まいびょう 推定 すいてい 値 ち 原因 げんいん 歩行 ほこう 痕跡 こんせき 数多 かずおお 発見 はっけん 一方 いっぽう 走行 そうこう 痕跡 こんせき 発見 はっけん 彼 かれ 走行 そうこう 示 しめ 可能 かのう 性 せい [ 61]
ティラノサウルス・レックスの後肢 あとあし 1993年 ねん 緩慢 かんまん 走行 そうこう 脚 あし 空中 くうちゅう 浮 う 存在 そんざい 主張 しゅちょう [ 62] 科 か 近 きん 縁 えん 動物 どうぶつ 最速 さいそく 大型 おおがた 獣 じゅう 脚 あし 類 るい 結論 けつろん [ 63] 脚 あし 骨 ほね 限定 げんてい 実際 じっさい 走行 そうこう 遥 はる 強靭 きょうじん 推定 すいてい 最 さい 高速度 こうそくど 約 やく 毎秒 まいびょう 提唱 ていしょう 速度 そくど 同 どう 程度 ていど [ 64] 達 たっ 高速 こうそく 運動 うんどう 中 ちゅう 転倒 てんとう 地面 じめん 叩 たた 付 つ 際 さい 小 ちい 前肢 ぜんし 衝撃 しょうげき 減衰 げんすい 重症 じゅうしょう 致命傷 ちめいしょう 負 お 主張 しゅちょう [ 65] [ 66] 動物 どうぶつ 園 えん 安全 あんぜん 環境 かんきょう 置 お キリン は骨折 こっせつ 危険 きけん 毎時 まいじ 行 おこな 知 し [ 67] [ 68] 必要 ひつよう 応 おう 高速 こうそく 動 うご 危険 きけん 受 う 入 い 可能 かのう 性 せい アロサウルス においても研究 けんきゅう [ 69] [ 70] 後 ご 研究 けんきゅう 走行 そうこう 速度 そくど 毎時 まいじ 毎時 まいじ 歩行 ほこう 緩慢 かんまん 走行 そうこう 中 なか 速 そく 走行 そうこう 以上 いじょう 絞 しぼ 込 こ [ 61] [ 71] [ 72] 化石 かせき 直接 ちょくせつ 得 え 用 もち 年 ねん 研究 けんきゅう 推定 すいてい 走行 そうこう 速度 そくど 毎秒 まいびょう [ 73] [ 74] 科 か 巨 きょ 大 だい 体重 たいじゅう 重 おも 運動 うんどう 能力 のうりょく 低 ひく 推測 すいそく 年 ねん 研究 けんきゅう 不利 ふり 補 おぎな 特徴 とくちょう 報告 ほうこく 一部 いちぶ 獣 しし 脚 あし 類 るい 見 み 顕著 けんちょ 卓越 たくえつ 尾大 びだい 腿 もも 筋 すじ 科 か 旧来 きゅうらい 復元 ふくげん 筋肉質 きんにくしつ 太 ふと 尾 お 部 ぶ 備 そな 尾大 びだい 腿 もも 筋 すじ 大腿 だいたい 骨 こつ 第 だい 四 よん 転 てん 子 こ 繋 つな 運動 うんどう 時 じ 脚 あし 部 ぶ 強力 きょうりょく 動 うご 可能 かのう 特 とく 他 た 科 か 尾大 びだい 腿 もも 筋 すじ 発達 はったつ [ 75]
Eric Snively らが2019年 ねん 発表 はっぴょう 研究 けんきゅう 科 か 同 どう アロサウルス上 うえ 科 か よりも機動 きどう 性 せい 高 たか 示唆 しさ 大型 おおがた 後肢 あとあし 筋肉 きんにく 結 むす 付 つ 体重 たいじゅう 比較 ひかく 回転 かいてん 慣性 かんせい 小 ちい 結果 けっか 科 か 相対 そうたい 的 てき 素早 すばや 旋回 せんかい 可能 かのう 獲物 えもの 近 ちか 際 さい 他 た 旋回 せんかい 際 さい 動作 どうさ 見 み 可能 かのう 性 せい 高 たか 旋回 せんかい 中 ちゅう 片 かた 脚 あし 固定 こてい 片方 かたがた 持 も 上 あ 動作 どうさ 可能 かのう 性 せい 仮説 かせつ 立 た 研究 けんきゅう 結果 けっか 類 るい 進化 しんか 敏捷 びんしょう 性 せい 寄与 きよ 過程 かてい 明 あき 可能 かのう 性 せい [ 76] 科 か 他 た 大型 おおがた 肉食 にくしょく 恐竜 きょうりゅう 見 み 特徴 とくちょう 的 てき 特異 とくい 的 てき 靱帯 じんたい 第 だい 三 さん 中 ちゅう 足 あし 骨 こつ 持 も 科 か 他 た 獣 しし 脚 あし 類 るい 体重 たいじゅう 重 おも 頑強 がんきょう 肉体 にくたい 一定 いってい 敏捷 びんしょう 性 せい 確保 かくほ 考 かんが [ 77]
加 くわ 年 ねん 研究 けんきゅう 科 か 例外 れいがい 的 てき 効率 こうりつ 的 てき 歩行 ほこう 者 しゃ 示唆 しさ et al. によると、ティラノサウルス科 か 含 ふく 種 しゅ 超 こ 獣 しし 脚 あし 類 るい 恐竜 きょうりゅう 後肢 あとあし 体重 たいじゅう 足取 あしど 比較 ひかく 調査 ちょうさ 様々 さまざま 方法 ほうほう 適用 てきよう 恐竜 きょうりゅう 最 さい 高速度 こうそくど 歩行 ほこう 時 じ 緩 ゆる 速度 そくど 運動 うんどう 時 じ 消費 しょうひ 量 りょう 推定 すいてい ドロマエオサウルス科 か などより小型 こがた 中型 ちゅうがた 種 たね 間 あいだ 長 なが 後肢 あとあし 速 はや 走行 そうこう 適応 てきおう 見 み 他 た 研究 けんきゅう 者 しゃ 以前 いぜん 結果 けっか 整合 せいごう 体重 たいじゅう 超 こ 獣 しし 脚 あし 類 るい 最高 さいこう 走行 そうこう 速度 そくど 体 からだ 制限 せいげん 受 う 長 なが 後肢 あとあし 速度 そくど 低 てい 歩行 ほこう 相関 そうかん 判明 はんめい 研究 けんきゅう 結果 けっか 小型 こがた 獣 しし 脚 あし 類 るい 進化 しんか 長 なが 脚 あし 狩 か 捕食 ほしょく 者 しゃ 逃走 とうそう 両方 りょうほう 役立 やくだ 一方 いっぽう 大型 おおがた 捕食 ほしょく 性 せい 獣 じゅう 脚 あし 類 るい 頂点 ちょうてん 捕食 ほしょく 者 しゃ 役割 やくわり 捕食 ほしょく 圧 あつ 解放 かいほう 進化 しんか 長 なが 脚 あし 減少 げんしょう 採 と 餌 えさ 効率 こうりつ 役立 やくだ 示唆 しさ 研究 けんきゅう 基盤 きばん 的 てき 獣 しし 脚 あし 類 るい 比較 ひかく 科 か 狩 か 死 し 肉 にく 漁 いさ 間 あいだ 支出 ししゅつ 減少 げんしょう 採 と 餌 えさ 効率 こうりつ 顕著 けんちょ 上昇 じょうしょう 示 しめ 結果 けっか 類 るい 彼 かれ 自身 じしん 維持 いじ 必要 ひつよう 狩 か 襲撃 しゅうげき 食餌 しょくじ 量 りょう 低減 ていげん 帰結 きけつ 可能 かのう 性 せい 高 たか 加 くわ 彼 かれ 研究 けんきゅう 類 るい 他 た 大型 おおがた 獣 じゅう 脚 あし 類 るい 俊敏 しゅんびん 示 しめ 研究 けんきゅう 繋 つな 中 なか 長距離 ちょうきょり 亘 わた 獲物 えもの 忍 しの 寄 よ 接近 せっきん 後 のち 突発 とっぱつ 的 てき 急 きゅう 以 もっ 殺害 さつがい 類 るい 長 た 示唆 しさ 結果 けっか 科 か 現生 げんなま オオカミ の間 あいだ 類似 るいじ 性 せい 指摘 してき 少 すく 一部 いちぶ 科 か 集団 しゅうだん 狩 か 行 おこな 裏付 うらづ [ 78] [ 79]
系統 けいとう 復元 ふくげん 古 こ 生物 せいぶつ 学 がく 続 つづ 議論 ぎろん 科 か 外皮 がいひ 被覆 ひふく 範囲 はんい 特性 とくせい 関 かん 前期 ぜんき 白 はく 亜紀 あき 義 ぎ 県 けん 累 るい 層 そう 中国 ちゅうごく 遼寧 りょうねい 省 しょう 分布 ぶんぷ 他 た 層 そう 無数 むすう 類 るい 骨格 こっかく 共 とも 長 なが タンパク質 たんぱくしつ 繊維 せんい 保存 ほぞん 産出 さんしゅつ [ 80] 他 た 仮説 かせつ 提唱 ていしょう [ 81] 通常 つうじょう 繊維 せんい 鳥類 ちょうるい 非 ひ 鳥類 ちょうるい 型 がた 獣 じゅう 脚 あし 類 るい 見 み 枝分 えだわ 羽毛 うもう 解釈 かいしゃく [ 82] [ 83] 年 ねん 記載 きさい ディロング の骨格 こっかく 含 ふく 上 うえ 科 か 確認 かくにん 最初 さいしょ 例 れい 現生 げんなま 鳥類 ちょうるい ダウンフェザー (英語 えいご 版 ばん 同様 どうよう 見 み 枝分 えだわ 大 だい 羽 はね (en にはなっておらず、体温 たいおん 保温 ほおん 用 もち 可能 かのう 性 せい [ 21] 年 ねん 全長 ぜんちょう 達 たっ ユウティラヌス が記載 きさい 本属 ほんぞく 発見 はっけん 大型 おおがた 獣 じゅう 脚 あし 類 るい 成体 せいたい 羽毛 うもう 生 は 可能 かのう 性 せい 示唆 しさ [ 84]
羽毛 うもう 生 は 彫像 ちょうぞう 系統 けいとう (英語 えいご 版 ばん 原則 げんそく 基 もと 科 か 羽毛 うもう 持 も 可能 かのう 性 せい 予想 よそう 年 ねん 研究 けんきゅう 州 しゅう 州 しゅう モンゴル国 こく で収集 しゅうしゅう 属 ぞく 科 か 皮膚 ひふ 印象 いんしょう 化石 かせき 記載 きさい 皮膚 ひふ 印象 いんしょう 化石 かせき 小型 こがた 頭部 とうぶ 後 こう 側 がわ 幅広 はばひろ 分布 ぶんぷ 腹部 ふくぶ 胸部 きょうぶ 腸 ちょう 骨 こつ 骨盤 こつばん 尾 お 部 ぶ 位置 いち 重複 じゅうふく 細 こま 小石 こいし 様 さま 密 みつ 鱗 うろこ 示 しめ 羽毛 うもう 痕跡 こんせき 無 な 基本 きほん 的 てき 直径 ちょっけい 約 やく 小 ちい 構成 こうせい 間 あいだ 約 やく 印象 いんしょう 散見 さんけん [ 85] 類 るい 足跡 あしあと 鱗 うろこ 見 み [ 86] 類 るい 顔面 がんめん 外皮 がいひ 歯 は 骨 こつ 上顎 じょうがく 骨 こつ 鱗 うろこ 表皮 ひょうひ 角質 かくしつ 化 か 下位 かい 領域 りょういき 鎧 よろい 皮膚 ひふ 研究 けんきゅう 判明 はんめい [ 87] [ 88]
Bell et al. はティラノサウルス上 うえ 科 か 外皮 がいひ 分布 ぶんぷ 既知 きち 内容 ないよう 基 もと 祖先 そせん 形質 けいしつ 復元 ふくげん 行 おこな 上 うえ 科 か 起源 きげん 羽毛 うもう 信頼 しんらい 性 せい 拘 かか 科 か 鱗 うろこ 持 も 信頼 しんらい 性 せい 真 しん 判断 はんだん 研究 けんきゅう 完全 かんぜん 扁平 へんぺい 上皮 じょうひ 覆 おお 説得 せっとく 力 りょく 証拠 しょうこ 皮膚 ひふ 印象 いんしょう 未 み 発見 はっけん 背中 せなか 側 がわ 羽毛 うもう 存在 そんざい 可能 かのう 性 せい 彼 かれ 認 みと et al. は、裏付 うらづ 根拠 こんきょ 必要 ひつよう 類 るい 鱗 うろこ 印象 いんしょう 羽毛 うもう 二 に 次 じ 的 てき 派生 はせい 可能 かのう 性 せい 仮説 かせつ 立 た [ 85] 科 か 起因 きいん 主張 しゅちょう [ 89]
そのような外皮 がいひ 何故 なぜ 発生 はっせい 決定 けってい 羽毛 うもう 喪失 そうしつ 前例 ぜんれい 鳥 とり 盤 ばん 類 るい 他 ほか 恐竜 きょうりゅう 見 み 過程 かてい 繊維状 せんいじょう 構造 こうぞう 失 うしな 鱗 うろこ 再 さい 出現 しゅつげん [ 90] 機 き 序 じょ 大型 おおがた 化 か 提唱 ていしょう 羽毛 うもう 体 からだ 近 ちか 指摘 してき 体 からだ 気候 きこう 類似 るいじ 挙 あ 差異 さい 理由 りゆう 分 わ [ 91]
ティラノサウルスの眼窩 がんか 前方 ぜんぽう 向 む 立体 りったい 視 し 可能 かのう ティラノサウルスは眼窩 がんか 前側 まえがわ 着 つ 眼球 がんきゅう 前側 まえがわ 向 む 現生 げんなま タカ よりも僅 わず 良 よ 両 りょう 眼 め 視 し 機能 きのう 持 も 一般 いっぱん 的 てき 捕食 ほしょく 性 せい 獣 じゅう 脚 あし 類 るい 頭蓋骨 ずがいこつ 真正面 ましょうめん 両 りょう 眼 め 視 し 機能 きのう 持 も 広範囲 こうはんい 視野 しや 重複 じゅうふく ジャック・ホーナー は、ティラノサウルス類 るい 系統 けいとう 着実 ちゃくじつ 両 りょう 眼 め 視 し 機能 きのう 向上 こうじょう 指摘 してき 類 るい 純粋 じゅんすい 腐肉 ふにく 食 しょく 動物 どうぶつ 場合 ばあい 立体 りったい 視 し 発達 はったつ 奥行 おくゆき 感覚 かんかく 不要 ふよう 長期 ちょうき 的 てき 傾向 けいこう 自然 しぜん 選択 せんたく 好 この 考 かんが [ 92] [ 93] 現生 げんなま 動物 どうぶつ 立体 りったい 視 し 主 おも 捕食 ほしょく 動物 どうぶつ 見 み 異 こと 他 た 科 か 典型 てんけい 的 てき 狭 せま 頭蓋骨 ずがいこつ 持 も 眼球 がんきゅう 主 おも 横 よこ 面 めん [ 94] 標本 ひょうほん 眼窩 がんか 楕円 だえん 形 がた 鍵穴 かぎあな 型 がた 円形 えんけい 他 た 科 か 属 ぞく 異 こと [ 12] 上下 じょうげ 高 たか 楕円 だえん 形 がた 見 み 円形 えんけい 見 み 鍵穴 かぎあな 型 がた 中 なか 間 あいだ [ 11] [ 12] [ 95]
ダスプレトサウルスと現生 げんなま 比較 ひかく 基 もと 年 ねん 詳細 しょうさい 研究 けんきゅう 科 か 大型 おおがた 平坦 へいたん 鱗 うろこ 持 も 判明 はんめい [ 96] [ 97] 鱗 うろこ 中心 ちゅうしん ケラチン 質 しつ 小型 こがた 存在 そんざい 化学 かがく 的 てき 刺激 しげき 熱 ねつ 的 てき 刺激 しげき 機械 きかい 的 てき 刺激 しげき 検知 けんち 可能 かのう 感覚 かんかく 束 たば 被覆 ひふく [ 98] [ 99] 彼 かれ 科 か 顔面 がんめん 鱗 うろこ 下 した 感覚 かんかく 束 たば 有 ゆう 科 か 用 もち 対象 たいしょう 特定 とくてい 巣 す 温度 おんど 計測 けいそく 静 しず 卵 たまご 幼体 ようたい 持 も 上 あ 可能 かのう 性 せい 提唱 ていしょう [ 96]
明瞭 めいりょう 鼻骨 びこつ 隆起 りゅうき 伴 ともな 頭蓋骨 ずがいこつ 数 すう 多 おお 科 か 恐竜 きょうりゅう 含 ふく 多数 たすう 獣 しし 脚 あし 類 るい 頭蓋骨 ずがいこつ 骨質 こっしつ 見 み 産出 さんしゅつ アリオラムス は、鼻骨 びこつ 上 じょう 卓越 たくえつ 個 こ 骨質 こっしつ 隆起 りゅうき 本 ほん 列 れつ 持 も 隆起 りゅうき 低 ひく 同様 どうよう 列 れつ アパタチオサウルス の頭蓋骨 ずがいこつ 存在 そんざい 一部 いちぶ 標本 ひょうほん 見 み [ 16] 涙 なみだ 骨 こつ 上 じょう 眼 め 正面 しょうめん 卓越 たくえつ 角 かく 存在 そんざい 涙 なみだ 骨 こつ 角 かく 存在 そんざい 代 か 後 こう 眼窩 がんか 骨 こつ 上 うえ 眼 め 後方 こうほう 三日月 みかづき 形 がた 存在 そんざい 頭部 とうぶ 用 もち 可能 かのう 性 せい 種 たね 認識 にんしき 求愛 きゅうあい 行動 こうどう 用 もち 可能 かのう 性 せい [ 11]
大 だい 部分 ぶぶん 恐竜 きょうりゅう 同様 どうよう 長 なが 外 そと 温 ゆたか 性 せい 考 かんが 年代 ねんだい 後半 こうはん 始 はじ 恐竜 きょうりゅう 初期 しょき ロバート・T・バッカー およびジョン・オストロム などの研究 けんきゅう 者 しゃ 見解 けんかい 対 たい 異議 いぎ 唱 とな [ 100] [ 101] 内 うち 温 ゆたか 性 せい 非常 ひじょう 活発 かっぱつ 生態 せいたい 示唆 しさ [ 102] 以降 いこう 複数 ふくすう 古 こ 生物 せいぶつ 学者 がくしゃ 体温 たいおん 調節 ちょうせつ 能力 のうりょく 明 あき 努 つと 若 わか 哺乳類 ほにゅうるい 鳥類 ちょうるい 比較 ひかく 高 たか 成長 せいちょう 率 りつ 持 も 的 てき 証拠 しょうこ 高 たか 代謝 たいしゃ 有 ゆう 仮説 かせつ 示唆 しさ 可能 かのう 性 せい 哺乳類 ほにゅうるい 鳥類 ちょうるい 同様 どうよう 成長 せいちょう 曲線 きょくせん 本 ほん 種 しゅ 成長 せいちょう 主 しゅ 未 み 成熟 せいじゅく 個体 こたい 制限 せいげん 示唆 しさ 永続 えいぞく 的 てき 成長 せいちょう 遂 と 他 ほか 脊椎動物 せきついどうぶつ 大半 たいはん 異 こと [ 45] 胴 どう 椎 しい 脛骨 けいこつ 体温 たいおん 差 さ 以下 いか 示唆 しさ 体 からだ 中枢 ちゅうすう 末梢 まっしょう 温度 おんど 範囲 はんい 小 ちい 古 こ 生物 せいぶつ 学者 がくしゃ 地球 ちきゅう 化学 かがく 者 しゃ 体内 たいない 温度 おんど 恒常 こうじょう 的 てき 外 そと 温 ゆたか 性 せい 爬虫類 はちゅうるい 内 うち 温 ゆたか 性 せい 哺乳類 ほにゅうるい 中 なか 間 あいだ 程度 ていど 代謝 たいしゃ 有 ゆう [ 103] 後 のち 数 すう 百 ひゃく 万 まん 年 ねん 過去 かこ 時代 じだい 別 べつ 大陸 たいりく 生息 せいそく ギガノトサウルス の標本 ひょうほん 彼 かれ 同様 どうよう 結果 けっか 発見 はっけん [ 104] 体温 たいおん 恒常 こうじょう 的 てき 維持 いじ 証拠 しょうこ 無 な 外 そと 温 ゆたか 性 せい 必 かなら 意味 いみ 体温 たいおん 調節 ちょうせつ 現生 げんなま ウミガメ のように、大型 おおがた 動物 どうぶつ 保温 ほおん 説明 せつめい [ 105] [ 106] [ 107]
シカゴのフィールド自然 しぜん 史 し 博物館 はくぶつかん にて、ダスプレトサウルスの骨格 こっかく ダイナソーパーク累 るい 層 そう 希少 きしょう 亜 あ 科 か 種 たね 共存 きょうぞん 属 ぞく 類 るい 共存 きょうぞん 数少 かずすく 例 れい 現生 げんなま 捕食 ほしょく 動物 どうぶつ 同様 どうよう 体 からだ 捕食 ほしょく 動物 どうぶつ 競争 きょうそう 制限 せいげん 解剖 かいぼう 学 がく 的 てき 行動 こうどう 学 がく 的 てき 地理 ちり 学 がく 的 てき 差異 さい 異 こと 生態 せいたい 的 てき 地位 ちい 分裂 ぶんれつ 累 るい 層 そう 科 か 分割 ぶんかつ 多 おお 不明 ふめい [ 108] 年 ねん 個体 こたい 数 すう 少 すく 狩 か 困難 こんなん 角 すみ 竜 りゅう 類 るい 曲 きょく 竜 りゅう 類 るい 残 のこ 仮説 かせつ 立 た [ 13] 累 るい 層 そう 産出 さんしゅつ 標本 ひょうほん 腹部 ふくぶ 消化 しょうか 類 るい 幼体 ようたい 遺骸 いがい 保存 ほぞん [ 109] 他 た 獣 しし 脚 あし 類 るい 異 こと 属 ぞく 標高 ひょうこう 差 さ 化石 かせき 産出 さんしゅつ 量 りょう 多寡 たか 認 みと [ 108] 累 るい 層 そう 北方 ほっぽう 層 そう 一般 いっぱん 的 てき 種 たね 南 みなみ 豊富 ほうふ 同様 どうよう 他 た 恐竜 きょうりゅう 見 み カンパニアン 期 き 北 きた 南西 なんせい カスモサウルス亜 あ 科 か およびハドロサウルス亜 あ 科 か 、北 きた 北部 ほくぶ セントロサウルス亜 あ 科 か およびランベオサウルス亜 あ 科 か )。ホルツは、このパターンはティラノサウルス亜 あ 科 か 亜 あ 科 か 亜 あ 科 か 間 あいだ 生態 せいたい 的 てき 嗜好 しこう 性 せい 共有 きょうゆう 示唆 しさ マーストリヒチアン 期 き 末 すえ 亜 あ 科 か エドモントサウルス のようなハドロサウルス亜 あ 科 か トリケラトプス のようなカスモサウルス亜 あ 科 か 北 きた 西部 せいぶ 広 ひろ 分布 ぶんぷ 一方 いっぽう 亜 あ 科 か 亜 あ 科 か 絶滅 ぜつめつ 亜 あ 科 か 衰退 すいたい [ 11]
ティラノサウルス科 か 間 あいだ 社会 しゃかい 的 てき 行動 こうどう 証拠 しょうこ 僅 わず 存在 そんざい 標本 ひょうほん 同 おな 産地 さんち 亜 あ 成体 せいたい 幼体 ようたい 骨格 こっかく 発見 はっけん 報告 ほうこく 類 るい 種 しゅ 社会 しゃかい 集団 しゅうだん 中 なか 生活 せいかつ 仮説 かせつ 支持 しじ 用 もち [ 110] 群居 ぐんきょ 性 せい 行動 こうどう 証拠 しょうこ 存在 そんざい [ 56] [ 57] 群 む 行動 こうどう 証拠 しょうこ 存在 そんざい [ 111]
ダイナソーパーク累 るい 層 そう 種 たね 若 わか 標本 ひょうほん 他 た 類 るい 起因 きいん 噛 か 後 ご 顔 かお 存在 そんざい 噛 か 跡 あと 治癒 ちゆ 噛 か 後 のち 個体 こたい 生 い 延 の 示唆 しさ 完全 かんぜん 成長 せいちょう 累 るい 層 そう 類 るい 噛 か 跡 あと 示 しめ 顔面 がんめん 攻撃 こうげき 若 わか 個体 こたい 限 かぎ 示唆 しさ 噛 か 後 ご 他 た 種 たね 可能 かのう 性 せい 顔面 がんめん 噛 か 付 つ 種 たね 内攻 ないこう 撃 げき 捕食 ほしょく 動物 どうぶつ 間 あいだ 非常 ひじょう 見 み 顔面 がんめん 噛 か 付 つ 他 ほか 類 るい シンラプトル やサウロルニトレステス といった他 ほか 獣 しし 脚 あし 類 るい 属 ぞく 見 み 噛 か 跡 あと 縄張 なわば 資源 しげん 社会 しゃかい 集団 しゅうだん 内 ない 支配 しはい 巡 めぐ 種 たね 内 ない 競争 きょうそう 起因 きいん 仮説 かせつ 立 た [ 56]
アルバートサウルスのジオラマ ダスプレトサウルスが社会 しゃかい 集団 しゅうだん 生息 せいそく 証拠 しょうこ 州 しゅう 累 るい 層 そう ボーンベッド から得 え 頭 とう 遺骸 いがい 含 ふく 大型 おおがた 成体 せいたい 小型 こがた 幼体 ようたい 中間 なかま 個体 こたい 同 どう 産地 さんち 少 すく 頭 とう 類 るい 保存 ほぞん 地質 ちしつ 学 がく 的 てき 証拠 しょうこ 遺骸 いがい 川 かわ 流 なが 集積 しゅうせき 全 すべ 動物 どうぶつ 同 おな 場所 ばしょ 同時 どうじ 埋没 まいぼつ 判明 はんめい 類 るい 遺骸 いがい 分散 ぶんさん 類 るい 歯 は 多数 たすう 噛 か 跡 あと 残 のこ 死亡 しぼう 時 じ 類 るい 採 と 餌 えさ 示唆 しさ 確証 かくしょう 持 も 群 む 形成 けいせい 推測 すいそく [ 57] 他 た 研究 けんきゅう 者 しゃ 他 た 大型 おおがた 獣 じゅう 脚 あし 類 るい 社会 しゃかい 集団 しゅうだん 形成 けいせい 証拠 しょうこ 懐疑 かいぎ 的 てき [ 112] 社会 しゃかい 相互 そうご 作用 さよう 現生 げんなま コモドオオトカゲ のものに類似 るいじ 群 む 非 ひ 協力 きょうりょく 的 てき 死骸 しがい 過程 かてい 互 たが 共食 ともぐ 頻繁 ひんぱん [ 113]
バーナム・ブラウンらが発見 はっけん 頭 とう 遺骸 いがい 保存 ほぞん 白 はく 亜紀 あき 獣 しし 脚 あし 類 るい 中 なか 最 もっと 多 おお 個体 こたい 場所 ばしょ 発見 はっけん 例 れい 大型 おおがた 獣 じゅう 脚 あし 類 るい 中 なか 州 しゅう 恐竜 きょうりゅう 採石 さいせき 場 じょう アロサウルス 群集 ぐんしゅう 次 つ 番目 ばんめ 多 おお 集団 しゅうだん 非常 ひじょう 高齢 こうれい 頭 とう 成体 せいたい 歳 さい 歳 さい 頭 とう 成体 せいたい 歳 さい 歳 さい 急 きゅう 成長 せいちょう 段階 だんかい 頭 とう 亜 あ 成体 せいたい 歳 さい 歳 さい 成長 せいちょう 段階 だんかい 達 たっ 頭 とう 幼体 ようたい 含 ふく [ 48] 植物 しょくぶつ 食 しょく 性 せい 動物 どうぶつ 遺骸 いがい 存在 そんざい 数 すう 多 おお 個体 こたい 保存 ほぞん 状態 じょうたい 類似 るいじ 産地 さんち カリフォルニア州 しゅう のラ・ブレア・タールピット のような捕食 ほしょく 動物 どうぶつ 罠 わな 保存 ほぞん 全 すべ 動物 どうぶつ 同時 どうじ 死亡 しぼう 結論 けつろん 群 む 行動 こうどう 根拠 こんきょ [ 114] 他 た 研究 けんきゅう 者 しゃ 懐疑 かいぎ 的 てき 旱魃 かんばつ 洪水 こうずい 他 た 要因 よういん 動物 どうぶつ 運搬 うんぱん 解釈 かいしゃく [ 48] [ 112] [ 115]
一般 いっぱん 議論 ぎろん 続 つづ 少 すく 科 か 種 たね 社会 しゃかい 的 てき 仮説 かせつ 支持 しじ 証拠 しょうこ 存在 そんざい ブリティッシュコロンビア州 しゅう のワピチ累 るい 層 そう 地元 じもと 住民 じゅうみん 頭 とう 科 か 個体 こたい 由来 ゆらい 足跡 あしあと 発見 はっけん 記載 きさい 足跡 あしあと 調査 ちょうさ 足跡 あしあと 作 つく 後 のち 長期間 ちょうきかん 放置 ほうち 痕跡 こんせき 認 みと 頭 とう 類 るい 個体 こたい 集団 しゅうだん 移動 いどう 仮説 かせつ 支持 しじ 調査 ちょうさ 動物 どうぶつ 時速 じそく 移動 いどう 腰 こし 高 たか 可能 かのう 性 せい 高 たか 判明 はんめい 層 そう 科 か 異 こと 属 ぞく 知 し 属 ぞく 足 あし 痕 こん 残 のこ 不明 ふめい [ 116] [ 117] [ 118] 証拠 しょうこ ユタ州 しゅう 南部 なんぶ 累 るい 層 そう 化石 かせき 証拠 しょうこ テラトフォネウス のものとされており、2021年 ねん 記載 きさい 他 た 科 か 社会 しゃかい 的 てき 動物 どうぶつ 示唆 しさ 歳 さい 亘 わた 異 こと 頭 とう 動物 どうぶつ 化石 かせき 洪水 こうずい 低 ひく 可能 かのう 性 せい 藍 あい 藻 も 毒 どく 火災 かさい 旱魃 かんばつ 引 ひ 起 お 大量 たいりょう 死 し 事変 じへん 示唆 しさ 属 ぞく 科 か 広 ひろ 社会 しゃかい 的 てき 行動 こうどう 見 み 意味 いみ 保存 ほぞん 全 すべ 動物 どうぶつ 短期間 たんきかん 死滅 しめつ 事実 じじつ 科 か 群居 ぐんきょ 性 せい 行動 こうどう 論拠 ろんきょ 補強 ほきょう [ 119] [ 120] [ 121]
ティラノサウルス類 るい 歯 は 型 がた 肉食 にくしょく 恐竜 きょうりゅう 食 しょく 痕 こん 最 もっと 広 ひろ 保存 ほぞん [ 122] 角 すみ 竜 りゅう 類 るい 類 るい 他 た 類 るい 報告 ほうこく [ 122] 既知 きち 科 か 骨 ほね 約 やく 歯 は 型 がた 保存 ほぞん [ 122] 科 か 歯 は ナイフ のように切断 せつだん 機能 きのう 肉 にく 体 からだ 引 ひ 剥 は 金具 かなぐ 用 もち [ 123] 歯 は 摩耗 まもう 類 るい 食餌 しょくじ 頭 あたま 振 ふ 複雑 ふくざつ 行動 こうどう 含 ふく 示唆 しさ [ 123]
パラサウロロフス を襲 おそ テラトフォネウス 少数 しょうすう 研究 けんきゅう 者 しゃ 群 む 狩猟 しゅりょう 行動 こうどう 群 む 若 わか 獲物 えもの 大型 おおがた 強力 きょうりょく 緩慢 かんまん 成体 せいたい 誘導 ゆうどう 推測 すいそく [ 114] 幼体 ようたい 成体 せいたい 異 こと 生態 せいたい 持 も 可能 かのう 性 せい 巨大 きょだい 成体 せいたい 小型 こがた 同 どう 時代 じだい 獣 しし 脚 あし 類 るい 間 あいだ 生 う 可能 かのう 性 せい 同 どう 時代 じだい 小型 こがた 獣 じゅう 脚 あし 類 るい 最大 さいだい 種 たね 成体 せいたい 体重 たいじゅう 桁 けた 小 ちい [ 11] 化石 かせき 記録 きろく 行動 こうどう 保存 ほぞん 非常 ひじょう 稀 まれ 容易 ようい 検証 けんしょう 可能 かのう 群 む 形成 けいせい 狩 か 行 おこな 思弁 しべん 確証 かくしょう 持 も 主張 しゅちょう 出来 でき [ 57] 行動 こうどう 証拠 しょうこ 発見 はっけん [ 56] [ 57]
ティラノサウルスが捕食 ほしょく 動物 どうぶつ 腐肉 ふにく 食 しょく 動物 どうぶつ 議論 ぎろん 関 かん 議論 ぎろん 同 どう 程度 ていど 昔 むかし 近 きん 縁 えん ゴルゴサウルス の骨格 こっかく 記載 きさい 歯 は 摩耗 まもう 基 もと 純粋 じゅんすい 腐肉 ふにく 食 しょく 動物 どうぶつ 結論 けつろん [ 124] 獣 しし 脚 あし 類 るい 歯 は 極 きわ 急速 きゅうそく 置換 ちかん 主張 しゅちょう 真剣 しんけん 受 う 止 と 最初 さいしょ 発見 はっけん 以降 いこう 大半 たいはん 研究 けんきゅう 者 しゃ 捕食 ほしょく 動物 どうぶつ 同意 どうい 現生 げんなま 捕食 ほしょく 動物 どうぶつ 機会 きかい 喜 よろこ 他 た 捕食 ほしょく 者 しゃ 獲物 えもの 盗 ぬす 死 し 肉 にく 漁 あさ 思 おも [ 125] [ 126]
ジャック・ホーナー は、ティラノサウルスが専 もっぱ 腐肉 ふにく 食 しょく 動物 どうぶつ 一切 いっさい 狩 か 行 おこな 考 かんが [ 62] [ 127] [ 128] 純粋 じゅんすい 腐肉 ふにく 食 しょく 動物 どうぶつ 仮説 かせつ 支持 しじ 複数 ふくすう 主張 しゅちょう 紹介 しょうかい 大型 おおがた 嗅球 と嗅神経 しんけい 存在 そんざい 遠距離 えんきょり 死骸 しがい 嗅 か 付 つ 高度 こうど 発達 はったつ 嗅覚 きゅうかく 示唆 しさ 歯 は 骨 ほね 砕 くだ 通常 つうじょう 最 もっと 栄養分 えいようぶん 少 すく 部位 ぶい 死体 したい 残 のこ 可能 かのう 限 かぎ 食料 しょくりょう 骨髄 こつづい 抜 ぬ 取 と 可能 かのう 高速 こうそく 走行 そうこう 苦手 にがて 早 はや 歩 ある 歩行 ほこう 主 おも 移動 いどう 手段 しゅだん 仮説 かせつ 一方 いっぽう 少 すく 獲物 えもの 動物 どうぶつ 俊敏 しゅんびん 動 うご 出来 でき [ 127] [ 129]
ゴルゴサウルスの幼体 ようたい 標本 ひょうほん TMP 2009.12.14 、シティペス (英語 えいご 版 ばん 化石 かせき 胃 い 中 なか 含 ふく 他 た 証拠 しょうこ 積極 せっきょく 的 てき 狩猟 しゅりょう 行動 こうどう 示唆 しさ 類 るい 眼窩 がんか 目 め 前 まえ 向 む 位置 いち 良好 りょうこう 立体 りったい 視 し 可能 かのう 類 るい 負 お 負傷 ふしょう トリケラトプス やハドロサウルス類 るい 骨格 こっかく 認 みと 最初 さいしょ 攻撃 こうげき 生 い 延 の 見 み [ 130] [ 131] [ 132] 腐肉 ふにく 食 しょく 動物 どうぶつ 北 きた 上部 じょうぶ 白亜 はくあ 系 けい 別 べつ 恐竜 きょうりゅう 頂点 ちょうてん 捕食 ほしょく 者 しゃ 地位 ちい 立 た 主張 しゅちょう 研究 けんきゅう 者 しゃ 頂点 ちょうてん 捕食 ほしょく 者 しゃ 獲物 えもの 大型 おおがた 周 しゅう 飾 かざり 頭 あたま 類 るい 鳥 とり 脚 あし 類 るい 他 た 科 か 数 すう 多 おお 特徴 とくちょう 共有 きょうゆう 除外 じょがい 場合 ばあい 頂点 ちょうてん 捕食 ほしょく 者 しゃ 候補 こうほ 残 のこ 小型 こがた ドロマエオサウルス類 るい のみである。このことから、腐肉 ふにく 食 しょく 動物 どうぶつ 仮説 かせつ 信 しん 者 もの 類 るい 巨体 きょたい 力 ちから 以 もっ 小型 こがた 動物 どうぶつ 獲物 えもの 盗 ぬす 可能 かのう 提唱 ていしょう [ 129]
ティラノサウルス科 か 少 すく 時折 ときおり 共食 ともぐ 強 つよ 示唆 しさ 証拠 しょうこ 自体 じたい 標本 ひょうほん 足 あし 上腕 じょうわん 骨 こつ 中 ちゅう 足 あし 骨 こつ 歯 は 型 がた 見 み 基 もと 少 すく 死骸 しがい 漁 あさ 共食 ともぐ 行 おこな 示唆 しさ [ 133] 層 そう 層 そう 共 とも 期 き 産 さん 化石 かせき 層 そう 期 き 産 さん 化石 かせき 盆地 ぼんち 様々 さまざま 科 か 共食 ともぐ 起 お 示唆 しさ 標本 ひょうほん 集 あつ 証拠 しょうこ 基 もと 科 か 同種 どうしゅ 動物 どうぶつ 共食 ともぐ 伴 ともな 日和見 ひよりみ 的 てき 摂食 せっしょく 行動 こうどう [ 134]
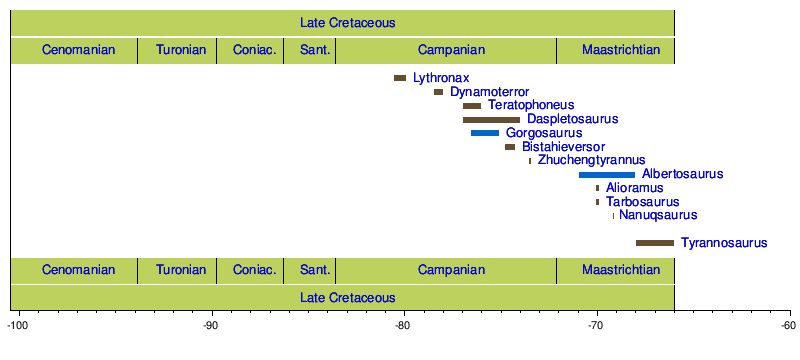
ティラノサウルス上 うえ 科 か 化石 かせき 分布 ぶんぷ 科 か 青色 あおいろ 示 しめ 初期 しょき 上 うえ 科 か 北半球 きたはんきゅう 全 すべ 大陸 たいりく 発見 はっけん 一方 いっぽう 科 か 化石 かせき 北 きた 発見 はっけん 南半球 みなみはんきゅう 産出 さんしゅつ 断片 だんぺん 的 てき 化石 かせき 科 か 報告 ほうこく 複 ふく 数 すう 回 かい アベリサウルス科 か の化石 かせき 誤 あやま 同定 どうてい [ 135] 白 はく 亜紀 あき 中頃 なかごろ 化石 かせき 記録 きろく 北 きた 両 りょう 地域 ちいき 乏 とぼ 科 か 起源 きげん 時代 じだい 場所 ばしょ 不明 ふめい 科 か 確認 かくにん 最初 さいしょ 期 き カンパニアン 期 き 北 きた 西部 せいぶ 生息 せいそく [ 11]
北 きた 東部 とうぶ 科 か 化石 かせき 発見 はっけん ドリプトサウルス やアパラチオサウルス といったより基盤 きばん 的 てき 上 うえ 科 か 化石 かせき 回収 かいしゅう 白 はく 亜紀 あき 終 お 持続 じぞく 白 はく 亜紀 あき 中頃 なかごろ 西部 せいぶ 内陸 ないりく 海路 かいろ 北 きた 分裂 ぶんれつ 後 のち 西部 せいぶ 科 か 進化 しんか 示唆 しさ [ 16] 科 か 化石 かせき アラスカ州 しゅう からも発見 はっけん 北 きた 分散 ぶんさん 可能 かのう 性 せい [ 136] 系統 けいとう 解析 かいせき 近 きん 縁 えん 位置 いち 置 お 本科 ほんか 固有 こゆう 系統 けいとう 形成 けいせい [ 18] 後 のち キアンゾウサウルス の発見 はっけん 族 ぞく 記載 きさい 反証 はんしょう [ 30] 年 ねん 日本 にっぽん 福井 ふくい 県立 けんりつ 恐竜 きょうりゅう 博物館 はくぶつかん 長崎 ながさき 県 けん 長崎 ながさき 市 し 長崎 ながさき 半島 はんとう 産出 さんしゅつ 科 か 歯 は 化石 かせき 報告 ほうこく 万 まん 年 ねん 前 まえ カンパニアン 期 き 日本 にっぽん 国内 こくない 初 はつ 大型 おおがた 類 るい 歯 は 化石 かせき 発表 はっぴょう [ 137]
タルボサウルスの骨格 こっかく ティラノサウルス亜 あ 科 か 広範囲 こうはんい 分布 ぶんぷ 亜 あ 科 か 発見 はっけん 族 ぞく 亜 あ 科 か 生息 せいそく 亜 あ 科 か 亜 あ 科 か 共 とも 期 き マーストリヒチアン 期 き 北 きた 生息 せいそく 亜 あ 科 か 西部 せいぶ 内陸 ないりく 中 ちゅう 分布 ぶんぷ 一方 いっぽう 亜 あ 科 か 大陸 たいりく 北西 ほくせい 部 ぶ 知 し [ 138]
後期 こうき 期 き 亜 あ 科 か 絶滅 ぜつめつ 亜 あ 科 か サスカチュワン州 しゅう からテキサス州 しゅう まで分布 ぶんぷ 他 た 北 きた 恐竜 きょうりゅう 当 あ 期 き 前期 ぜんき 期 き 北西 ほくせい 部 ぶ ランベオサウルス亜 あ 科 か の鳥 とり 脚 あし 類 るい セントロサウルス亜 あ 科 か の角 すみ 竜 りゅう 繁栄 はんえい 南部 なんぶ ハドロサウルス亜 あ 科 か とカスモサウルス亜 あ 科 か が繁栄 はんえい 白 はく 亜紀 あき 末 まつ 亜 あ 科 か 発見 はっけん 亜 あ 科 か 数 かず 減 へ 一方 いっぽう 亜 あ 科 か 亜 あ 科 か 西部 せいぶ 内陸 ないりく 中 ちゅう 繁栄 はんえい [ 11] 年 ねん 北 きた 西部 せいぶ 大 だい 部分 ぶぶん 他 ほか 科 か 絶滅 ぜつめつ 自体 じたい 部分 ぶぶん 的 てき 関与 かんよ 示唆 しさ 研究 けんきゅう 北 きた 進化 しんか 子孫 しそん 渡 わた 他 た 科 か 競合 きょうごう 生存 せいぞん 競争 きょうそう 勝利 しょうり 示唆 しさ 仮説 かせつ 知 し 範囲 はんい 他 た 種 しゅ 科 か 発見 はっけん 事実 じじつ 支持 しじ [ 139]
^ 全長 ぜんちょう 標本 ひょうほん 番号 ばんごう FMNH PR 2081愛称 あいしょう 体重 たいじゅう 標本 ひょうほん 番号 ばんごう RSM P2523.8 愛称 あいしょう 別 べつ 個体 こたい 推定 すいてい
^ Holtz, Thomas R. Jr. (2012) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix. Archived 2017-08-12 at the Wayback Machine .
^ a b グレゴリー・ポール (著 ちょ 恐竜 きょうりゅう 事典 じてん 原著 げんちょ 第 だい 版 はん 東洋一 とよかず 今井 いまい 拓哉 たくや 河 かわ 部 ぶ 壮 たけし 一郎 いちろう 柴田 しばた 正輝 まさき 関谷 せきや 透 とおる 服部 はっとり 創 はじめ 紀 きの 訳 わけ 共立 きょうりつ 出版 しゅっぱん 年 ねん 月 がつ 日 にち 頁 ぺーじ ISBN 978-4-320-04738-9
^ a b c Hutchinson, John R.; Bates, Karl T.; Molnar, Julia; Allen, Vivian; Makovicky, Peter J.; Claessens, Leon (2011年 ねん "A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny, and Growth" . PLOS ONE . 6 (10): e26037. Bibcode :2011PLoSO...626037H . doi :10.1371/journal.pone.0026037 PMC 3192160 PMID 22022500 。
^ Therrien, F.; Henderson, D. M. (2007年 ねん Journal of Vertebrate Paleontology . 27 (1): 108–115. doi :10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . ISSN 0272-4634 . S2CID 86025320 。 ^ Hartman, Scott (July 7, 2013). “Mass estimates: North vs South redux ”. Scott Hartman's Skeletal Drawing.com. October 12, 2013時 じ 点 てん オリジナル よりアーカイブ。August 24, 2013 閲覧 えつらん ^ a b c d e Carpenter, Ken. (1992年 ねん Aspects of Nonmarine Cretaceous Geology . Beijing: China Ocean Press. pp. 250–268.
^ Breithaupt, B.H.; Southwell, E.H.; Matthews, N.A. (18 October 2005). "In Celebration of 100 years of Tyrannosaurus rex : Manospondylus gigas , Ornithomimus grandis , and Dynamosaurus imperiosus , the Earliest Discoveries of Tyrannosaurus rex in the West" . Abstracts with Programs . 2005 Salt Lake City Annual Meeting . Vol. 37. Geological Society of America. p. 406. 2019年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Breithaup, BH; Southwell EH; Matthews NA (2006年 ねん Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus rex in Wyoming and the West". New Mexico Museum of Natural History and Science Bulletin . 35 : 257–258. ^ Bakker R.T; Williams M.; Currie P. (1988年 ねん Nanotyrannus , a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana". Hunteria . 1 : 1–30. ^ a b Kurzanov, Sergei M. "A new carnosaur from the Late Cretaceous of Nogon-Tsav, Mongolia". The Joint Soviet-Mongolian Paleontological Expedition Transactions (ロシア語 ご 3 : 93–104.
^ a b c d e f g h i j k l m n o p q r Holtz, Thomas R. (2004年 ねん The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 111–136. ISBN 978-0-520-24209-8
^ a b c d e f g Currie, Philip J. (2003年 ねん "Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta" (PDF) . Acta Palaeontologica Polonica . 48 (2): 191–226. 2018年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。2010年 ねん 月 がつ 日 にち 閲覧 えつらん 。
^ a b c d Russell, Dale A. (1970年 ねん National Museum of Natural Sciences Publications in Paleontology . 1 : 1–34.
^ Maleev, Evgeny A. (1955年 ねん "New carnivorous dinosaurs from the Upper Cretaceous of Mongolia" (PDF) . Doklady Akademii Nauk SSSR (ロシア語 ご 104 (5): 779–783. 2012年 ねん 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。2008年 ねん 月 がつ 10日 とおか 閲覧 えつらん 。 ^ Currie, Philip J. (2000年 ねん The Age of Dinosaurs in Russia and Mongolia . Cambridge: Cambridge University Press. pp. 434–455. ISBN 978-0-521-54582-2 ^ a b c d e f Carr, Thomas D.; Williamson, Thomas E.; Schwimmer, David R. (2005年 ねん Journal of Vertebrate Paleontology . 25 (1): 119–143. doi :10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2 . ISSN 0272-4634 . S2CID 86243316 。
^ Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (2017年 ねん 月 がつ 日 にち "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system" . Scientific Reports (英語 えいご 7 (1): 44942. Bibcode :2017NatSR...744942C . doi :10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 PMID 28358353 。 ^ a b c d Currie, Philip J.; Hurum, Jørn H; Sabath, Karol. (2003年 ねん "Skull structure and evolution in tyrannosaurid phylogeny" (PDF) . Acta Palaeontologica Polonica . 48 (2): 227–234. 2008年 ねん 月 がつ 日 にち 時点 じてん アーカイブ (PDF) 。2010年 ねん 月 がつ 日 にち 閲覧 えつらん 。
^ a b c d e f g Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84–89. In: Mesozoic Vertebrate Life . Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
^ Quinlan, Elizabeth D.; Derstler, Kraig; Miller, Mercedes M. (2007年 ねん "Anatomy and function of digit III of the Tyrannosaurus rex manus" . Geological Society of America Annual Meeting — Abstracts with Programs : 77. 2008年 ねん 月 がつ 日 にち 時点 じてん アーカイブ 。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ a b c Xu Xing, X ; Norell, Mark A.; Kuang Xuewen; Wang Xiaolin; Zhao Qi; Jia Chengkai. (2004年 ねん Nature . 431 (7009): 680–684. Bibcode :2004Natur.431..680X . doi :10.1038/nature02855 . PMID 15470426 . S2CID 4381777 。
^ Holtz, Thomas R. (1994年 ねん Journal of Paleontology . 68 (5): 1100–1117. doi :10.1017/S0022336000026706 . JSTOR 1306180 . S2CID 129684676 。 ^ Cope E.D. (1866年 ねん Proceedings of the Academy of Natural Sciences of Philadelphia . 18 : 275–279. ^ Maleev E.A. (1955年 ねん Doklady Akademii Nauk SSSR . 104 (5): 779–783. ^ Leidy, Joseph (1856年 ねん Proceedings of the Academy of Natural Sciences of Philadelphia . 8 : 72–73.^ Osborn, Henry F. (1905年 ねん Tyrannosaurus and other Cretaceous carnivorous dinosaurs". Bulletin of the American Museum of Natural History . 21 (3): 259–265. doi :10.1111/j.1468-5965.2007.00735_17.x . hdl :2246/1464 。^ Liddell, Henry G.; Scott, Robert (1980年 ねん Greek-English Lexicon ISBN 978-0-19-910207-5 ^ Fiorillo, A. R.; Tykoski, R. S. (2014年 ねん Dodson, Peter (ed.). "A Diminutive New Tyrannosaur from the Top of the World" . PLoS ONE 9 (3): e91287. Bibcode :2014PLoSO...991287F . doi :10.1371/journal.pone.0091287 PMC 3951350 PMID 24621577 。 ^ Paul, Gregory S. (1988年 ねん Predatory Dinosaurs of the World ISBN 978-0-671-61946-6 ^ a b Lü, Junchang; Yi, Laiping; Brusatte, Stephen L.; Yang, Ling; Li, Hua; Chen, Liu (2014年 ねん "A new clade of Asian Late Cretaceous long-snouted tyrannosaurids" . Nature Communications . 5 : 3788. Bibcode :2014NatCo...5.3788L . doi :10.1038/ncomms4788 PMID 24807588 。
^ “Pinocchio rex long-snouted tyrannosaur discovered in Asia ” (7 May 2014). 2 February 2019時 じ 点 てん アーカイブ 。8 May 2014 閲覧 えつらん ^ Olshevsky, George (1995年 ねん Kyoryugaku Saizensen [Dino Frontline] . 9–10 : 92–119. ^ Olshevsky G.; Ford T.L. (1995年 ねん Dino Frontline . 6 : 75–99. ^ Fowler, DW; Woodward, HN; Freedman, EA; Larson, PL; Horner, JR (2011年 ねん "Reanalysis of "Raptorex kriegsteini": A Juvenile Tyrannosaurid Dinosaur from Mongolia" . PLOS ONE . 6 (6): e21376. Bibcode :2011PLoSO...6E1376F . doi :10.1371/journal.pone.0021376 PMC 3126816 PMID 21738646 。 ^ Sereno, Paul C. (1998年 ねん Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen (ドイツ語 ご 210 : 41–83. doi :10.1127/njgpa/210/1998/41 。 ^ a b c d e f g h Holtz, TR, Jr. (2001), The phylogeny and taxonomy of the Tyrannosauridae in K Carpenter & D Tanke [eds.], Mesozoic Vertebrate Life. Indiana Univ. Press, pp. 64-83.
^ Brochu, Christopher R. (2003年 ねん Tyrannosaurus rex : insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull". Society of Vertebrate Paleontology Memoirs . 7 : 1–138. doi :10.2307/3889334 . JSTOR 3889334 。 ^ Sereno, Paul C. (7 November 2005). “Stem Archosauria—TaxonSearch ”. 19 February 2007時 じ 点 てん オリジナル よりアーカイブ。14 January 2008 閲覧 えつらん ^ Hurum, Jørn H.; Sabath, Karol. (2003年 ねん "Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared" (abstract) . Acta Palaeontologica Polonica . 48 (2): 161–190. 2012年 ねん 月 がつ 日 にち 時点 じてん アーカイブ 。2008年 ねん 月 がつ 10日 とおか 閲覧 えつらん 。 ^ “Newly found dinosaur is long-nosed cousin of Tyrannosaurus rex ”. ScienceDaily . 2018年 ねん 月 がつ 日 にち 時点 じてん アーカイブ 。2018年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Loewen, M.A.; Irmis, R.B.; Sertich, J.J.W.; Currie, P. J. ; Sampson, S. D. (2013年 ねん Evans, David C (ed.). "Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans" . PLoS ONE 8 (11): e79420. Bibcode :2013PLoSO...879420L . doi :10.1371/journal.pone.0079420 PMC 3819173 PMID 24223179 。 ^ a b c d e f g h Erickson, Gregory M., GM; Makovicky, Peter J.; フィリップ・J・カリー; Norell, Mark A.; Yerby, Scott A.; Brochu, Christopher A. (2004年 ねん Nature . 430 (7001): 772–775. Bibcode :2004Natur.430..772E . doi :10.1038/nature02699 . PMID 15306807 . S2CID 4404887 . Sampled longevities for T. rex ranged from 2 to 28 years
^ a b Horner, J.R. & Padian, K. (2004年 ねん "Age and growth dynamics of Tyrannosaurus rex" . Proceedings of the Royal Society of London B . 271 (1551): 1875–1880. doi :10.1098/rspb.2004.2829 . PMC 1691809 PMID 15347508 。
^ Lee, Andrew H.; Werning, Sarah (2008年 ねん "Sexual maturity in growing dinosaurs does not fit reptilian growth models" . Proceedings of the National Academy of Sciences . 105 (2): 582–587. Bibcode :2008PNAS..105..582L . doi :10.1073/pnas.0708903105 PMC 2206579 PMID 18195356 。 ^ Schweitzer, M.H., Wittmeyer, J.L., & Horner, J.R. (2005年 ねん "Gender-specific reproductive tissue in ratites and Tyrannosaurus rex " . Science . 308 (5727): 1456–1460. Bibcode :2005Sci...308.1456S . doi :10.1126/science.1112158 . PMID 15933198 . S2CID 30264554 . 2021年 ねん 月 がつ 10日 とおか 時点 じてん オリジナル よりアーカイブ。2019年 ねん 日 にち 閲覧 えつらん 。 ^ a b c d e Erickson, G.M., Currie, P.J., Inouye, B.D., & Winn, A.A. (2006年 ねん "Tyrannosaur life tables: an example of nonavian dinosaur population biology" . Science . 313 (5784): 213–217. Bibcode :2006Sci...313..213E . doi :10.1126/science.1125721 . PMID 16840697 . S2CID 34191607 . 2021年 ねん 月 がつ 10日 とおか 時点 じてん オリジナル よりアーカイブ。2019年 ねん 日 にち 閲覧 えつらん 。
^ “First tyrannosaur embryo fossils revealed ”. Science (October 19, 2020). December 5, 2020時 じ 点 てん オリジナル よりアーカイブ。November 23, 2020 閲覧 えつらん ^ October 2020, Laura Geggel-Associate Editor 15 (15 October 2020). “1st of their kind baby tyrannosaur fossils unearthed ”. livescience.com . 17 October 2020時 じ 点 てん オリジナル よりアーカイブ。17 October 2020 閲覧 えつらん ^ “Fat-footed tyrannosaur parents couldn't keep up with their skinnier offspring, fossil footprints reveal ”. The Conversation . 2021年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2021年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Enriquez, Nathan J.; Campione, Nicolás E.; Brougham, Tom; Fanti, Federico; White, Matt A.; Sissons, Robin L.; Sullivan, Corwin; Vavrek, Matthew J.; Bell, Phil R. (2021年 ねん 月 がつ 日 にち "Exploring possible ontogenetic trajectories in tyrannosaurids using tracks from the Wapiti Formation (upper Campanian) of Alberta, Canada" . Journal of Vertebrate Paleontology . 40 (6): e1878201. doi :10.1080/02724634.2021.1878201 ^ Erickson, Gregory M.; Curry Rogers, Kristi; Varricchio, David J.; Norell, Mark; Xu, Xing (2007年 ねん "Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition" . Biology Letters . 3 (5): 558–561. doi :10.1098/rsbl.2007.0254 . PMC 2396186 PMID 17638674 。 ^ a b c Lee, Andrew H.; Werning, Sarah (2008年 ねん "Sexual maturity in growing dinosaurs does not fit reptilian growth models" . PNAS . 105 (2): 582–587. Bibcode :2008PNAS..105..582L . doi :10.1073/pnas.0708903105 PMC 2206579 PMID 18195356 。
^ Ricklefs, Robert E. (2007年 ねん "Tyrannosaur ageing" . Biology Letters . 3 (2): 214–217. doi :10.1098/rsbl.2006.0597 . PMC 2375931 PMID 17284406 。 ^ a b c d Tanke, Darren H.; Currie, Philip J. (1998年 ねん "Head-biting behavior in theropod dinosaurs: paleopathological evidence" (PDF) . Gaia . 15 : 167–184. 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。
^ a b c d e Currie, Philip J.; Trexler, David; Koppelhus, Eva B.; Wicks, Kelly; Murphy, Nate (2005年 ねん The Carnivorous Dinosaurs . Bloomington: Indiana University Press. pp. 313–324. ISBN 978-0-253-34539-4
^ “Tyrannosaurus had poor turning circle ”. May 7, 2012時 じ 点 てん オリジナル よりアーカイブ。2007年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Carrier, D.R., Walter, R.M., and Lee, D.V. (2001年 ねん 日 にち "Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia" . Journal of Experimental Biology . 204 (Pt 22): 3917–3926. doi :10.1242/jeb.204.22.3917 . PMID 11807109 . 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Snively, Eric; O’Brien, Haley; Henderson, Donald M.; Mallison, Heinrich; Surring, Lara A.; Burns, Michael E.; Jr, Thomas R. Holtz; Russell, Anthony P.; Witmer, Lawrence M. (2019年 ねん 月 がつ 日 にち "Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods" . PeerJ (英語 えいご 7 : e6432. doi :10.7717/peerj.6432 . ISSN 2167-8359 。 ^ a b Hutchinson, J.R. (2004年 ねん "Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa" (PDF) . Journal of Morphology . 262 (1): 441–461. doi :10.1002/jmor.10240 . PMID 15352202 . S2CID 15677774 . 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。
^ a b Horner, John R.; Don Lessem (1993年 ねん The complete T. rex ISBN 978-0-671-74185-3
^ Holtz, Thomas R. (1996年 ねん 月 がつ 日 にち "Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda)" . Journal of Paleontology . 70 (3): 536–538. doi :10.1017/S0022336000038506 . S2CID 87599102 . 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Christiansen, P. (1998年 ねん "Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential" (PDF) . Gaia . 15 : 241–255. ISSN 0871-5424 . 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Farlow, James O. ; Smith, Matthew B.; Robinson, John M. (1995年 ねん "Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex " . Journal of Vertebrate Paleontology . 15 (4): 713–725. doi :10.1080/02724634.1995.10011257 . 2008年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。^ "The bigger they come, the harder they fall" Archived 2015-05-31 at the Wayback Machine . New Scientist, 7 October 1995, p. 18.^ “Giraffe ”. WildlifeSafari.info. 12 June 2010時 じ 点 てん アーカイブ 。29 April 2006 閲覧 えつらん ^ “The History of Woodland Park Zoo — Chapter 4 ”. June 2, 2007時 じ 点 てん オリジナル よりアーカイブ。29 April 2006 閲覧 えつらん ^ Alexander, R.M. (2006年 ねん 月 がつ 日 にち "Dinosaur biomechanics" . Proc Biol Sci . 273 (1596): 1849–1855. doi :10.1098/rspb.2006.3532 . PMC 1634776 PMID 16822743 。 ^ Hanna, Rebecca R. (2002年 ねん Allosaurus fragilis ) with comparisons to allosaur pathology in the Cleveland-Lloyd Dinosaur Quarry collection". Journal of Vertebrate Paleontology . 22 (1): 76–90. doi :10.1671/0272-4634(2002)022[0076:MIAIIA]2.0.CO;2 . ISSN 0272-4634 . S2CID 85654858 。 ^ Hutchinson, John R.; Garcia, M (2002年 ねん 月 がつ 日 にち Tyrannosaurus was not a fast runner". Nature . 415 (6875): 1018–1021. Bibcode :2002Natur.415.1018H . doi :10.1038/4151018a . PMID 11875567 . S2CID 4389633 。 ^ Hajdul, R. (1997). Tendons. Dinosaur Cards . Orbis Publishing Ltd. D36044311.
^ Sellers, W.I. & Manning, P.L. (2007年 ねん 月 がつ "Estimating dinosaur maximum running speeds using evolutionary robotics" . Proc. R. Soc. B . 274 (1626): 2711–6. doi :10.1098/rspb.2007.0846 . PMC 2279215 PMID 17711833 . 2011年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Liz Seward: "T. rex 'would outrun footballer'" Archived 2012-01-07 at the Wayback Machine .. BBC News website, Tuesday, 21 August 2007. The article quotes Dr Bill Sellers, University of Manchester, co-author of a paper published in Proceedings of the Royal Society B . Retrieved 22 August 2007.^ Persons, W. Scott; Currie, Philip J. (2010年 ねん 日 にち "The Tail of Tyrannosaurus: Reassessing the Size and Locomotive Importance of the M. caudofemoralis in Non-Avian Theropods" . The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology . 294 (1): 119–131. doi :10.1002/ar.21290 . ISSN 1932-8486 。 ^ Snively, Eric; O’Brien, Haley; Henderson, Donald M.; Mallison, Heinrich; Surring, Lara A.; Burns, Michael E.; Jr, Thomas R. Holtz; Russell, Anthony P.; Witmer, Lawrence M.; Currie, Philip J.; Hartman, Scott A.; Cotton, John R. (2019年 ねん 月 がつ 日 にち "Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods" . PeerJ . 7 : e6432. doi :10.7717/peerj.6432 PMC 6387760 PMID 30809441 。 ^ Surring, Lara; Burns, Michael; Snively, Eric; Barta, Daniel; Holtz, Thomas; Russell, Anthony; Witmer, Lawrence; Currie, Philip (2022年 ねん 日 にち "Consilient evidence affirms expansive stabilizing ligaments in the tyrannosaurid foot" . Vertebrate Anatomy Morphology Palaeontology (英語 えいご 10 (1). doi :10.18435/vamp29387 . ISSN 2292-1389 。 ^ Dececchi, T. Alexander; Mloszewska, Aleksandra M.; Holtz, Thomas R.; Habib, Michael B.; Larsson, Hans C. E. (2020年 ねん "The fast and the frugal: Divergent locomotory strategies drive limb lengthening in theropod dinosaurs" . PLOS ONE . 15 (5): e0223698. Bibcode :2020PLoSO..1523698D . doi :10.1371/journal.pone.0223698 PMC 7220109 PMID 32401793 。 ^ “T. rex was a champion walker, super-efficient at lower speeds ”. EurekAlert! . 2020年 ねん 月 がつ 日 にち 時点 じてん アーカイブ 。2020年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Zhou Zhonghe, Z; Barrett, Paul M.; Hilton, Jason. (2003年 ねん Nature . 421 (6925): 807–814. Bibcode :2003Natur.421..807Z . doi :10.1038/nature01420 . PMID 12594504 . S2CID 4412725 。 ^ Lingham-Soliar, Theagarten; Feduccia, Alan; Wang, Xiaolin (2007年 ねん "A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres" . Proceedings of the Royal Society of London. Series B, Biological Sciences . 274 (1620): 1823–1829. doi :10.1098/rspb.2007.0352 . PMC 2270928 PMID 17521978 。 ^ Chen Peiji, Pei-ji; Dong Zhiming ; Zhen Shuonan. (1998年 ねん Nature . 391 (6663): 147–152. Bibcode :1998Natur.391..147C . doi :10.1038/34356 . S2CID 4430927 。 ^ Xu Xing, X ; Zhou Zhonghe; Prum, Richard A. (2003年 ねん Nature . 410 (6825): 200–204. Bibcode :2001Natur.410..200X . doi :10.1038/35065589 . PMID 11242078 . S2CID 4426803 。^ “Archived copy ”. 2012年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2012年 ねん 月 がつ 日 にち 閲覧 えつらん ^ a b Bell, P. R.; Campione, N. E.; Persons, W. S.; Currie, P. J.; Larson, P. L.; Tanke, D. H.; Bakker, R. T. (2017年 ねん "Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution" . Biology Letters . 13 (6): 20170092. doi :10.1098/rsbl.2017.0092 PMC 5493735 PMID 28592520 。
^ Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003. The First Late Cretaceous Footprints from the Nemegt Locality in the Gobi of Mongolia. Ichnos. Vol.10:1-12.Currie, P.J., Badamgarav, D., Koppelhus, E.B. 2003.
^ Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M., & Moore, J. R. (2017). A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Scientific Reports , 7.
^ “MORPHOLOGY, TAXONOMY, AND PHYLOGENETIC RELATIONSHIPS OF THE MONTEVIALE CROCODYLIANS (OLIGOCENE, ITALY). ”. The Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy (September 2018). hdl :/2318/1703198 2024年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Saitta, Evan T.; Fletcher, Ian; Martin, Peter; Pittman, Michael; Kaye, Thomas G.; True, Lawrence D.; Norell, Mark A.; Abbott, Geoffrey D.; Summons, Roger E.; Penkman, Kirsty; Vinther, Jakob (2018年 ねん 日 にち "Preservation of feather fibers from the Late Cretaceous dinosaur Shuvuuia deserti raises concern about immunohistochemical analyses on fossils" (PDF) . Organic Geochemistry (英語 えいご 125 : 142–151. doi :10.1016/j.orggeochem.2018.09.008 . ISSN 0146-6380 . S2CID 105753275 。 ^ blogger, Guest. “Ten Things We Don't Know about Tyrannosaurs ”. 2020年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2021年 ねん 月 がつ 日 にち 閲覧 えつらん ^ “Tyrannosaurus rex had scaly skin and wasn’t covered in feathers, a new study says ”. The Washington Post (2017年 ねん 月 がつ 日 にち 年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2022年 ねん 月 がつ 10日 とおか 閲覧 えつらん ^ Stevens, Kent A. (2006年 ねん 月 がつ "Binocular vision in theropod dinosaurs" (PDF) . Journal of Vertebrate Paleontology . 26 (2): 321–330. doi :10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2 . ISSN 0272-4634 . S2CID 85694979 . 2012年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。 ^ Jaffe, Eric (2006年 ねん 月 がつ 日 にち "Sight for 'Saur Eyes: T. rex vision was among nature's best" . Science News . 170 (1): 3–4. doi :10.2307/4017288 . JSTOR 4017288 . 2012年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Saveliev, Sergei V.; Alifanov, Vladimir R. (2005年 ねん Tarbosaurus bataar (Theropoda, Tyrannosauridae)". Paleontological Journal . 41 (3): 281–289. doi :10.1134/S0031030107030070 . S2CID 53529148 。 ^ Carr, Thomas D. (1999年 ねん "Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria)" . Journal of Vertebrate Paleontology . 19 (3): 497–520. doi :10.1080/02724634.1999.10011161 . S2CID 83744433 . 2020年 ねん 月 がつ 20日 はつか 時点 じてん アーカイブ 。2019年 ねん 日 にち 閲覧 えつらん 。 ^ a b Carr, Thomas D.; Varricchio, David J.; Sedlmayr, Jayc C.; Roberts, Eric M.; Moore, Jason R. (2017年 ねん 月 がつ 日 にち "A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system" . Scientific Reports (英語 えいご 7 : 44942. Bibcode :2017NatSR...744942C . doi :10.1038/srep44942 . ISSN 2045-2322 . PMC 5372470 PMID 28358353 。
^ Barker, Chris; Naish, Darren; Katsamenis, Orestis; Dyke, Gareth (2017年 ねん 月 がつ 日 にち "Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii" . Scientific Reports . 7 (1): 3749. Bibcode :2017NatSR...7.3749B . doi :10.1038/s41598-017-03671-3 PMC 5473926 PMID 28623335 。 ^ Leitch, Duncan B.; Catania, Kenneth C. (2012年 ねん 日 にち "Structure, innervation and response properties of integumentary sensory organs in crocodilians" . Journal of Experimental Biology (英語 えいご 215 (23): 4217–4230. doi :10.1242/jeb.076836 . ISSN 0022-0949 . PMC 4074209 PMID 23136155 。 ^ Di-Poï, Nicolas; Milinkovitch, Michel C. (2013年 ねん 月 がつ 日 にち "Crocodylians evolved scattered multi-sensory micro-organs" . EvoDevo . 4 (1): 19. doi :10.1186/2041-9139-4-19 . ISSN 2041-9139 . PMC 3711810 PMID 23819918 。 ^ Bakker, Robert T. (1968年 ねん Discovery . 3 (2): 11–12. ^ Bakker, Robert T. (1972年 ねん Nature . 238 (5359): 81–85. Bibcode :1972Natur.238...81B . doi :10.1038/238081a0 . S2CID 4176132 。 ^ Bakker, Robert T. (1986年 ねん The Dinosaur Heresies ISBN 978-0-688-04287-5 OCLC 13699558 。 [要 よう 番号 ばんごう ^ Barrick, Reese E.; William J. Showers (1994年 ねん 月 がつ Science . 265 (5169): 222–224. Bibcode :1994Sci...265..222B . doi :10.1126/science.265.5169.222 . PMID 17750663 . S2CID 39392327 。 ^ Barrick, Reese E.; William J. Showers (1999年 ねん 月 がつ "Thermophysiology and biology of Giganotosaurus : comparison with Tyrannosaurus " . Palaeontologia Electronica . 2 (2). 2011年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Paladino, Frank V.; Spotila, James R.; Dodson, Peter (1999年 ねん The Complete Dinosaur . Bloomington: Indiana University Press. pp. 491–504. ISBN 978-0-253-21313-6 ^ Chinsamy, Anusuya; Willem J. Hillenius (2004年 ねん The dinosauria . Berkeley: University of California Press. pp. 643–659. ISBN 978-0-520-24209-8 ^ Seymour, Roger S. (2013年 ねん 月 がつ 日 にち "Maximal Aerobic and Anaerobic Power Generation in Large Crocodiles versus Mammals: Implications for Dinosaur Gigantothermy" . PLOS ONE . 8 (7): e69361. Bibcode :2013PLoSO...869361S . doi :10.1371/journal.pone.0069361 ISSN 1932-6203 . PMC 3702618 PMID 23861968 。 ^ a b Farlow, James O.; Pianka, Eric R. (2002年 ねん "Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs" . Historical Biology . 16 (1): 21–40. doi :10.1080/0891296031000154687 . S2CID 18114585 . 2021年 ねん 月 がつ 10日 とおか 時点 じてん オリジナル よりアーカイブ。2019年 ねん 日 にち 閲覧 えつらん 。
^ Varricchio, David J. (2001年 ねん Journal of Paleontology . 75 (2): 401–406. doi :10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2 . ISSN 0022-3360 . S2CID 86113170 。 ^ Guinness World Records Ltd. (2003). 2003 Guinness World Records . pg 90.
^ Strganac, C., Jacobs L., Polcyn M., Mateus O., Myers T., Araújo R., Fergunson K. M., Gonçalves A. O., Morais M. L., Schulp A. S., da Tavares T. S., & Salminen J. (2014). Geological Setting and Paleoecology of the Upper Cretaceous Bench 19 Marine Vertebrate Bonebed at Bentiaba, Angola. Netherlands Journal of Geosciences. 1-16.
^ a b Eberth, David A.; McCrea, Richard T. (2001年 ねん Journal of Vertebrate Paleontology . 21 (Supplement to Number 3): 46A. doi :10.1080/02724634.2001.10010852 . S2CID 220414868 。
^ Roach, Brian T.; Brinkman, Daniel L. (2007年 ねん Deinonychus antirrhopus and other nonavian theropod dinosaurs". Bulletin of the Peabody Museum of Natural History . 48 (1): 103–138. doi :10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 。 ^ a b Currie, Philip J. (1998年 ねん "Possible evidence of gregarious behavior in tyrannosaurids" (PDF) . Gaia . 15 : 271–277. 2009年 ねん 月 がつ 日 にち 時点 じてん オリジナル (PDF) よりアーカイブ。2008年 ねん 月 がつ 日 にち 閲覧 えつらん 。
^ Roach, Brian T.; Brinkman, Daniel T. (2007年 ねん Deinonychus antirrhopus and other nonavian theropod dinosaurs". Bulletin of the Peabody Museum of Natural History . 48 (1): 103–138. doi :10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2 . S2CID 84175628 。 ^ McCrea, Richard T.; Buckley, Lisa G.; Farlow, James O.; Lockley, Martin G.; Currie, Philip J.; Matthews, Neffra A.; Pemberton, S. George (2014年 ねん "A 'Terror of Tyrannosaurs': The First Trackways of Tyrannosaurids and Evidence of Gregariousness and Pathology in Tyrannosauridae" . PLOS ONE . 9 (7): e103613. Bibcode :2014PLoSO...9j3613M . doi :10.1371/journal.pone.0103613 PMC 4108409 PMID 25054328 。 ^ “Tyrannosaurus Tracks Show Dinosaur's Walk ” (2014年 ねん 月 がつ 日 にち 年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2015年 ねん 月 がつ 日 にち 閲覧 えつらん ^ “Tracks Hint at the Social Life of Tyrant Dinosaurs ” (2014年 ねん 月 がつ 日 にち 年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2015年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Titus, Alan L.; Knoll, Katja; Sertich, Joseph J. W.; Yamamura, Daigo; Suarez, Celina A.; Glasspool, Ian J.; Ginouves, Jonathan E.; Lukacic, Abigail K.; Roberts, Eric M. (2021年 ねん 月 がつ 日 にち "Geology and taphonomy of a unique tyrannosaurid bonebed from the upper Campanian Kaiparowits Formation of southern Utah: implications for tyrannosaurid gregariousness" . PeerJ . 9 : e11013. doi :10.7717/peerj.11013 PMC 8061582 PMID 33976955 。 ^ Eilperin, Juliet (April 19, 2021). “Tyrannosaurs likely hunted in packs rather than heading out solo, scientists find” . The Washington Post . オリジナル のApril 20, 2021時点 じてん . https://archive.today/20210420094932/https://www.washingtonpost.com/climate-environment/2021/04/19/grand-staircase-tyrannosaur/ September 27, 2021 閲覧 えつらん ^ “Tyrannosaurus Rex 'hunted in packs' ”. 2021年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2021年 ねん 月 がつ 日 にち 閲覧 えつらん ^ a b c Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
^ a b Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozoic Vertebrate Life . Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
^ Lambe, L. B. (1917年 ねん "The Cretaceous theropodous dinosaur Gorgosaurus " . Memoirs of the Geological Survey of Canada . 100 : 1–84. doi :10.4095/101672 . 2021年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2021年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Kowalewski, M.: “The Fossil Record of Predation ”. pp. 251–266 (2002年 ねん 年 ねん 月 がつ 日 にち 時点 じてん オリジナル よりアーカイブ。2022年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Dorey, M. (1997). Tyrannosaurus. Dinosaur Cards . Orbis Publishing Ltd. D36045907.
^ a b Horner, J.R. (1994年 ねん Tyrannosaurus as a scavenger)". The Paleontological Society Special Publication . 7 : 157–164. doi :10.1017/S2475262200009497 。
^ Amos, J. (31 July 2003). “T. rex goes on trial” . BBC. オリジナル の13 March 2012時点 じてん . https://web.archive.org/web/20120313130020/http://news.bbc.co.uk/1/hi/sci/tech/3112527.stm 12 October 2008 閲覧 えつらん ^ a b Walters, M., Paker, J. (1995). Dictionary of Prehistoric Life. Claremont Books. ISBN 1-85471-648-4 .
^ Erickson G. M.; Olson K. H. (1996年 ねん "Bite marks attributable to Tyrannosaurus rex : preliminary description and implications" . Journal of Vertebrate Paleontology . 16 (1): 175–178. doi :10.1080/02724634.1996.10011297 . 2021年 ねん 月 がつ 日 にち 時点 じてん アーカイブ 。2020年 ねん 月 がつ 日 にち 閲覧 えつらん 。 ^ Carpenter, K. (1998年 ねん "Evidence of predatory behavior by theropod dinosaurs" . Gaia . 15 : 135–144. 2007年 ねん 日 にち 時点 じてん オリジナル よりアーカイブ。2007年 ねん 日 にち 閲覧 えつらん 。 ^ Fowler, Denver W.; Robert M. Sullivan (2006年 ねん New Mexico Museum of Natural History and Science Bulletin . 35 : 127–130. ^ Longrich, N. R.; Horner, J. R.; Erickson, G. M.; Currie, P. J. (2010年 ねん "Cannibalism in Tyrannosaurus rex " . PLOS ONE . 5 (10): e13419. Bibcode :2010PLoSO...513419L . doi :10.1371/journal.pone.0013419 PMC 2955550 PMID 20976177 。 ^ “NEW EVIDENCE FOR CANNIBALISM IN TYRANNOSAURID DINOSAURS FROM THE UPPER CRETACEOUS (CAMPANIAN/MAASTRICHTIAN) SAN JUAN BASIN OF NEW MEXICO ”. ResearchGate . 2021年 ねん 月 がつ 10日 とおか 時点 じてん オリジナル よりアーカイブ。2021年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Dodson, Peter; Britt, Brooks; Carpenter, Kenneth; Forster, Catherine A.; Gillette, David D.; Norell, Mark A.; Olshevsky, George; Parrish, J. Michael & Weishampel, David B. (1993年 ねん Age of Dinosaurs . Publications International, LTD. p. 105. ISBN 978-0-7853-0443-2 ^ Fiorillo, Anthony R.; Gangloff, Roland A. (2000年 ねん Journal of Vertebrate Paleontology . 20 (4): 675–682. doi :10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2 . ISSN 0272-4634 . S2CID 130766946 。 ^ “国内 こくない 初 はつ 科 か 大型 おおがた 種 しゅ 歯 は 化石 かせき 福井 ふくい 県立 けんりつ 恐竜 きょうりゅう 博物館 はくぶつかん 年 ねん 月 がつ 日 にち 2022年 ねん 月 がつ 日 にち 閲覧 えつらん ^ Thomas R. Holtz Jr. (1994年 ねん Journal of Paleontology . 68 (5): 1100–1117. doi :10.1017/S0022336000026706 . JSTOR 1306180 . S2CID 129684676 。 ^ February 2016, Laura Geggel 29 (29 February 2016). “T. Rex Was Likely an Invasive Species ”. livescience.com . 14 July 2019時 じ 点 てん アーカイブ 。5 January 2021 閲覧 えつらん







_of_Daspletosaurus_horneri.jpg)





















.jpg)












